阅读:0
听报道
有些行为是天生的,有些可为后天改变;
动物求偶是天生的,经历可变求偶强弱。
谁求谁?一般来说,对后代投入较少(或者不投入)的性别,追求产后对后代投入较多的性别。虽然多数动物是雄性追求雌性,也有例外:少数动物雄性对后代关爱很多,那些种动物的雌性就比较在意追求雄性。
有追求,就有竞争,也有失败,动物也会失恋。
对雄性来说,失恋对个体可能不是好事,但可能对群体是好事,也可能说明雌性比较“忠贞”。我们可以看看老鼠和果蝇的不同情形:在实验室常用的老鼠中(不是所有种系),雌老鼠与雄老鼠交配、而怀孕后,在一些天内,雌鼠如果遇到第二只雄鼠,雌鼠会自动流产,再和第二只雄鼠交配。而实验室常用的果蝇不是这样。雌蝇与雄蝇交配后,如果再来第二只雄蝇,无论他多么热情、多么健壮、多么…,她拒绝第三者插足,不仅逃开、还用后腿踢他。进化中为什么出现较“忠贞”雌性的果蝇、较“随意”型雌性的老鼠,是研究进化的生物学家的问题,而神经生物学家关注的是机理。
对群体是好事,对具体的雄性就不一定。比如,捷足先登的雄果蝇首先交配雌果蝇后,可以放心大胆地离开去干其他事情。而如果是第二只雄蝇,不知道雌蝇已经交配、或者知道但没有其他机会,不幸追交配过的雌蝇,而被名花有主的雌蝇不客气地反复拒绝、甚至暴力教训后,那就有点惨了。饱受挫折后,如果再遇到挺乐意被追的处女果蝇,他也打不起精神来,不愿追雌性。雄果蝇的这种行为变化,是Brandeis大学的Siegel和Hall于1979年发现的,学名为求偶条件化(courtship conditioning),也称为求偶学习(courtship learning),当然这是负学习,雄蝇天生碰到雌蝇就会求偶,而被拒绝后学会少求偶、不求偶。
雌蝇为什么交配后拒绝雄蝇?是她有忠贞不移的天性?还是第一只雄蝇的计谋?前些年的研究表明,第一只成功交配雌蝇的雄蝇在将生殖细胞送给雌蝇的同时,液体中夹带了一个蛋白质分子,它刺激雌蝇的神经系统,导致雌蝇以后不愿交配。
第二只倒霉的雄蝇失恋以后,如何学得不追雌蝇?前人的研究表明,第一只交配成功的雄蝇,在交配时还干过第二件事,他将自己体表的一种雄性特有的小分子cVA粘到雌蝇体表,导致第二只雄蝇倒胃口,cVA本身抑制雄蝇交配。第一只雄蝇双管齐下、内外夹攻,改变了雌蝇、也影响以后以第三者身份试图交配的其他雄蝇。
失恋的雄蝇脑如何学得不追雌蝇?Brandeis大学生化系主任Griffith等实验室的几个实验表明,雄蝇脑内一个称为蘑菇体的结构起作用。因为研究果蝇的人多年来已知蘑菇体对嗅觉记忆非常重要,而已知挥发性分子cVA刺激嗅觉,所以发现蘑菇体参与求偶学习,较易理解。
我实验室最近研究雄蝇求偶学习的机理,乃从另一研究穿插过来。我们研究动物的社会行为,包括果蝇、老鼠和人。清华本科毕业、通过科学院生物物理所加入我实验室的研究生周传,研究了神经递质鱆胺在果蝇打架中起重要作用,他为第一作者于2008年发表的Nature Neuroscience论文表明,如果没有鱆胺,雄蝇对打降低、雌蝇对打也降低,所以,鱆胺不分性别在两性中都参与打架行为。鱆胺是递质分子,从一个神经细胞分泌后,作用于下一个细胞膜表面的受体。周传希望找到鱆胺下游的受体分子中哪个参与打架。果蝇有四个鱆胺受体,周传首先选了1998年被韩裔美国人发现的OAMB。结果他发现OAMB不参与打架,但我们已对OAMB做了一些相关工作,为了不浪费这些努力,周传寻找OAMB还可能起的作用,试了以后发现,OAMB参与求偶学习,这样阴差阳错地,我实验室一位研究生开始和其他实验室合作研究了果蝇失恋的分子机理。
周传发现,鱆胺对于求偶学习是必需的。如果缺失TbH基因,雄蝇就不能合成鱆胺。这样的雄蝇在追求已交配雌蝇失败后,不能学会减少求偶,如果给他一只处女蝇,他像没事似的追。如果用转基因把合成酶的基因导入突变雄蝇,他追已交配雌蝇后沮丧地不追处女蝇,也就是有正常的求偶学习。如果通过遗传学方法短暂地抑制鱆胺能神经细胞释放递质分子,也可以使雄蝇不能求偶学习。缺失基因的实验,雄蝇一生都缺乏TbH,不能确定基因什么时候影响行为。而抑制递质释放的实验,可以很好地控制时间(精确到数秒),这样周传发现鱆胺能神经细胞只在求偶学习的学习期起作用。就是说,在已交配雌蝇教训雄蝇时,需要雄蝇脑内的鱆胺能神经细胞开展工作,才能使雄蝇以后减少求偶。
雄蝇希望交配雌蝇,是受雌蝇的吸引。雄蝇受教训后不追处女蝇,是因为不受处女吸引吗?周传发现不是,因为缺乏TbH基因的雄蝇照常被雌蝇吸引,只是不能接受已交配雌蝇的教训。因此,周传假设鱆胺起的作用是介导雄蝇涂在雌蝇身上的cVA之作用。为此,他用遗传操纵激活鱆胺能神经元,结果可以抑制雄蝇对处女蝇的追求。而且,如果不用已交配雌蝇,仅用处女蝇,但同时激活鱆胺能神经元,这样雄蝇并未被雌蝇教训,结果经历这样过程的雄蝇事后也不追求处女蝇,表明激活鱆胺能神经元能模拟cVA的作用,这些结果显示鱆胺能神经元的作用是介导cVA的信号。
前人用原位杂交发现鱆胺受体OAMB的mRNA表达在蘑菇体,参与嗅觉学习记忆。
我们与科学院生物物理所焦仁杰和他的学生黄海合作,他们将酵母转录因子Gal4引入果蝇OAMB基因中,这样Gal4模拟OAMB的表达模式。他们获得这个转基因品系的果蝇后,我们观察发现OAMB-Gal4确实表达在蘑菇体中。
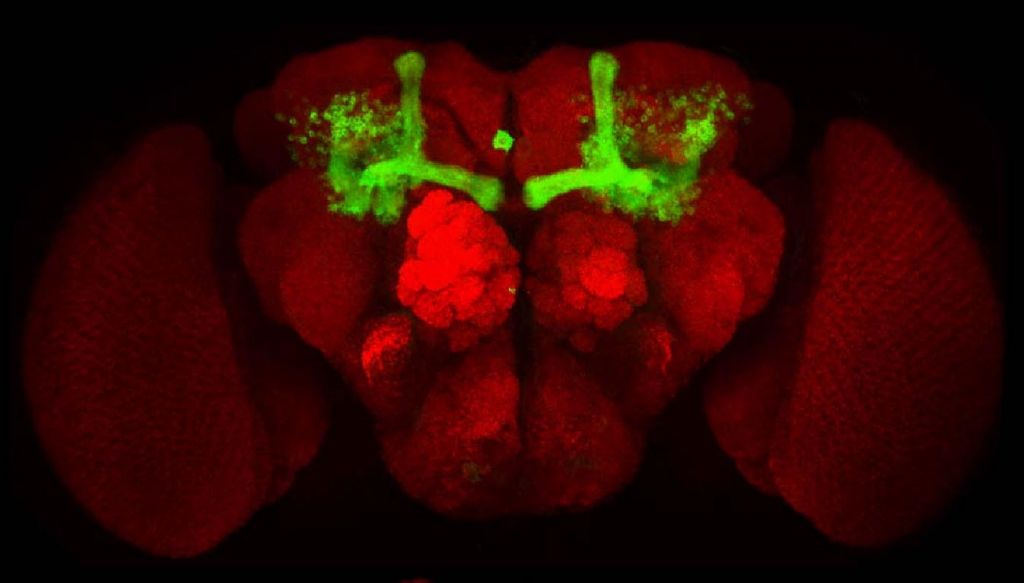
用OAMB-Gal4,我们可以控制其他基因的表达,比如具有抑制和激活神经元活性的基因,让它们表达到含OAMB的神经细胞,然后操纵这些细胞的活性,从而研究含OAMB神经细胞的功能。周传发现,短暂抑制OAMB神经细胞,可以使雄蝇不能在接触已交配雌蝇后被教训后识相,说明OAMB神经细胞对求偶学习是必需的。用OAMB基因突变的雄蝇做实验,验证了OAMB基因本身对求偶学习也是必需的。
我们前面介绍了,鱆胺能神经细胞不仅对于求偶学习是必需的,而且是充分的,充分性是通过激活鱆胺能神经细胞来验证的。那么,OAMB是否对求偶学习是充分的,也可以通过激活OAMB神经细胞来检验。周传做了这个实验,发现OAMB 神经细胞对于求偶学习不是充分的。而且,激活OAMB神经细胞不能抑制雄蝇对处女蝇的追求。所以,应该还有其他鱆胺受体,而不是OAMB,介导鱆胺的这些作用。
如果鱆胺和OAMB介导cVA的作用,那么,含OAMB的神经细胞就应该对cVA有反应。神经细胞的反应性,目前有一种检测方法是看细胞内钙浓度的改变。周传到美国开会时,事先联系了UCSD王竞实验室,到他们那里学习做果蝇神经细胞钙成像,他也把OAMB-Gal4果蝇送到美国了。他开完会后,与王竞实验室的Kim合作。周传看到体外给cVA可以导致雄蝇OAMB神经细胞内钙浓度升高。他回国后,请Kim在UCSD继续和重复此实验,开始她没有看到,后来重复了,并有比较好的照片,发表为我们文章的图4。不过,除OAMB神经细胞以外可能还有对cVA反应的神经细胞。我们实验室后来也有了必要的成像系统、建立了钙成像的检测方法。
我们希望看鱆胺能神经细胞与含OAMB的神经细胞是否有连接,这个实验是通过在前一种细胞表达半个分子,后一种细胞表达半个分子,如果这两半分开,看不到荧光,如果两半在物理距离上很接近,可以发出荧光。在台湾的清华大学江安世实验室有很好的工具,我们与他的研究生林暄文合作,周传把我们的果蝇寄去,他们将合适的对象合起来,结果发现鱆胺能神经细胞确实与蘑菇体的神经细胞有近距离接触,一般认为这表明有突触连接。江安世实验室拍的很漂亮的图片是我们论文的图5。含OAMB的神经细胞位于蘑菇体,所以,以上实验间接推测鱆胺能神经细胞与OAMB神经细胞的关系,而尚无实验直接确定它们的关系。
求偶学习实际是一种条件反射(conditioned reflex)。我们知道,俄国生理学家巴普洛夫(Ivan Pavlov,1849-1936)在因为用狗研究消化生理学而获1904年诺贝尔奖时,已专注于研究他通过用唾液和胃液而发现的条件反射。他发现,给狗食物的时候,还没有入口,狗就开始流唾液,这是狗天生就会的自然行为。而如果在提供食物时,同时给另外一个原本与食物无关、单独不会诱导流唾液的信号,比如铃声,将铃声与食物在时间上同步,多次重复,久而久之,只给铃声也能诱导狗流唾液。也就是说,经过训练,狗学会铃声与食物的相关性,以为铃声来了食物也会来,所以对铃声的反应相同于食物:产生更多唾液,为吃食物做准备。在这个实验中,食物称为非条件刺激,因为无需其他条件,先天就能导致固定的反应,而铃声称为条件刺激,唾液腺对铃声原本无反应,是在铃声与食物时间相关的条件下成为对唾液的刺激。
雄蝇的求偶学习也可以从条件反射来理解。雄蝇碰到雌蝇本不会放弃求偶,而雄蝇碰到雄蝇分泌的cVA会减少求偶,cVA作为非条件刺激。因为交配过的雌蝇身上带有前面雄蝇涂抹的cVA,cVA与雌蝇身体相关,雌蝇(可能是其体表特异的外激素)变成了条件刺激,经过一段时间,雄蝇以为有雌蝇气味的都不是好惹的,所以失去追求雌性的愿望。
周传等认为,鱆胺在cVA下游工作,在脑内介导cVA这一非条件刺激的作用。他在没有cVA的情况下,给雄蝇提供处女雌蝇时,同时激活鱆胺能神经细胞,结果使雄蝇学会回避雌蝇,支持鱆胺是在cVA下游的可能,他和Kim通过钙成像观察到cVA可以激活OAMB神经细胞的结果进一步支持这个可能。不过,并未完全证明,因缺乏直接观察鱆胺能神经细胞对cVA的反应,而OAMB本身的激活不足以代替cVA起到非条件刺激的作用(教训雄蝇不追雌蝇)。所以,我们的工作还有不完善和不清楚的地方。不完善的那部分,是我们觉得既然OAMB已对cVA有反应,推测鱆胺能神经细胞对cVA的反应问题不大,所以没做。不清楚的是既然激活OAMB不能完全模拟cVA、也不能模拟鱆胺能神经细胞的非条件刺激作用,那么就有其他鱆胺受体起这样的作用,是哪个、或哪几个鱆胺受体介导非条件刺激的作用,尚需研究。
此研究中我们发现鱆胺可以参加惩罚性刺激。几年前,德国的Heisenberg实验室提出果蝇中,两种神经递质(鱆胺和多巴胺)分别对应于奖赏性刺激和惩罚性刺激,这个假设不为我们的研究结果所支持。在哺乳类动物,多巴胺对奖赏很重要,平时起奖赏作用,而过分的、不合适的奖赏就是成瘾和赌博等的基础。现在看来,似乎果蝇中并非简单的一种递质参与奖赏、一种参与惩罚。
2012年10月发表于J Neuroscience (《神经科学杂志》)的文章,题目是“性拒绝的分子遗传学分析:鱆胺及其受体OAMB参与果蝇求偶条件化”揭示了果蝇脑内鱆胺在求偶学习中必需和充分的作用,OAMB受体为求偶学习所必需、但不充分;鱆胺能神经细胞与蘑菇体的神经细胞有突触连接;还有其他受体参与介导鱆胺的作用。
文章作者包括:周传、黄海、Suzy Kim、林暄文、孟祥兰,合作实验室是科学院生物物理所的焦仁杰、UCSD王竞、台湾清华大学的江安世、美国德州大学的Han。
此课题是我们研究果蝇GPCR作用的一部分,因为OAMB和其他鱆胺受体,都是G蛋白偶联受体(GPCR)。在北京进行的研究获得科技部和北京市的支持。
我们研究鱆胺原本是关注打架的分子机理。这条线索并未结束,虽然OAMB受体不参与打架,其他受体应该参与。
注1:提出果蝇鱆胺和多巴胺对应奖赏和惩罚关系的德国神经生物学家Martin Heisenberg(1940-),乃26岁提出测不准原理的物理学家Werner Heisenberg(海森堡,1901-1976)之子。Martin之兄Jochen Heisenberg子继父业为物理学家。而Martin于1966年从德国Tübingen大学获生物化学博士学位,到加州理工学院随德裔物理学家转成生物学家的Max Delbrück做博士后,当时加州理工的Seymour Benzer开始用果蝇研究神经生物学,Martin Heisenberg自1968年一直用果蝇遗传学研究行为的神经生物学,特别是果蝇的学习记忆。他哥哥在美国任教,他在德国任教。
注2:以前我实验室最多和一、两个实验室合作,周传等这篇文章创了我实验室合作实验室数量的记录(有五个)。这么多实验室列在文章中,出现过地址问题。投稿过程中,我被告知台湾“国科会”不允许作者用China作为地址,允许用Taiwan、或“Taiwan, Republic of China”。我不懂为什么台湾规定可以用Republic of China,而不能用去政治化的、更为简单的China。我就此事询问台湾“国科会”主任、副主任,他们不回答我的email、却捅到台湾新闻媒体,我只好在博客上回应。因为台湾科学家不按规定怕失去科研经费支持,在我不理解其规则缘由的情况下,我们最后只好两种表述:台湾作者地址是“Taiwan 30013,Republic of China”,北大作者地址是“Beijing 100871, People’s Republic of China”。
进一步阅读
Siegel RW, Hall JC (1979) Conditioned responses in courtship behavior of normal and mutant Drosophila. Proc Natl Acad Sci USA 76:3430 –3434.
Ejima A, Smith BP, Lucas C, Levine JD, Griffith LC (2005) Sequential learning of pheromonal cues modulates memory consolidation in trainerspecific associative courtship conditioning. Curr Biol 15:194 –206.
Ejima A, Smith BP, Lucas C, van der Goes van Naters W, Miller CJ, Carlson JR, Levine JD, Griffith LC (2007) Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate. Curr Biol 17:599–605.
Gailey DA, Jackson FR, Siegel RW (1984) Conditioning mutations in Drosophila melanogaster affect an experience-dependent behavioral modification in courting males. Genetics 106:613– 623.
Han KA, Millar NS, Davis RL (1998) A novel octopamine receptor with preferential expression in Drosophila mushroom bodies. J Neurosci 18:3650–3658.
McBride SM, Giuliani G, Choi C, Krause P, Correale D, Watson K, Baker G, Siwicki KK (1999) Mushroom body ablation impairs short-term memory and long-term memory of courtship conditioning in Drosophila melanogaster. Neuron 24:967–977.
Schwaerzel M, Monastirioti M, Scholz H, Friggi-Grelin F, Birman S, Heisenberg M (2003) Dopamine and octopamine differentiate between aversive and appetitive olfactory memories in Drosophila. J Neurosci 23:10495–10502.
Lee HG, Seong CS, Kim YC, Davis RL, HanKA (2003) Octopamine receptor OAMB is required for ovulation in Drosophila melanogaster. Dev Biol 264:179 –190.
Siwicki KK, Riccio P, Ladewski L, Marcillac F, Dartevelle L, Cross SA, Ferveur JF (2005) The role of cuticular pheromones in courtship conditioning of Drosophila males. Learn Mem 12:636–645.
Zhou C, Rao Y, Rao Y (2008) A subset of octopaminergic neurons are important for Drosophila aggression. Nat Neurosci 11:1059 –1067.
Zhou C, Huang H, Kim SM, Lin H, Meng X, Han KA, Chiang A-S, Wang JW, Jiao R, Rao Y (2012) Molecular genetic analysis of sexual rejection: roles of octopamine and its receptor OAMB in Drosophila courtship conditioning. J Neurosci 32:14281–14287.
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}