基础研究通过推进人类认知前沿有时可解决人类面临的问题;人类面临的问题通过激发科学研究有时也推进人类认识前沿。遗传的科学问题与肺炎的医疗问题合力推动揭示了基因的物质基础。
科学前沿可以出现不同研究途径的意外交汇。从生物学功能的角度诞生了遗传学:1866年Mendel开创遗传学,1880年德国的Flemming发现染色体,1902年德国的Boveri获得证据支持染色体为遗传的物质基础,1910年后美国的Morgan及其学生丰富和发展了染色体的遗传学说。而从生物的化学成分之角度出现了生物化学:1869年,瑞士的Miescher在研究伤口脓细胞化学成分的过程中发现核素,其后德国的Kossel发现了核酸中的嘌呤和嘧啶,二十世纪初美国的Levene确定以核苷酸连接为基础的核酸一级结构,瑞典的Caspersson等证明染色体含核酸和蛋白质,但当时研究DNA生物化学和生物物理的专家以为DNA无特异性、缺乏信息携带能力。
1928年,研究肺炎致病性细菌的过程中,英国的Griffith分析不同类型病例分布后推测不同型的肺炎球菌可能会变化,之后设计实验发现了转化现象。1944年,美国洛克菲勒医学研究所的Avery、MacLeod和McCarty研究转化的物质基础,提出脱氧核糖核酸(DNA)是改变细菌可遗传特性的转化因子。其后研究验证DNA的转化活性、证明DNA有特异性、发现转化活性不局限于特定细菌的特定性状。
DNA是遗传的物质基础的概念刺激进一步研究,最重要的是1953年Watson和Crick在Franklin和Wilkins对DNA进行X线衍射的基础上提出DNA结构的双螺旋模型,分子生物学随之而诞生。
1核酸及其化学结构
1-1 核酸的发现
米歇尔(Johann Friedrich Miescher,1844-1895)出生于科学世家,父亲曾任瑞士的巴塞尔(Basel)大学生理学教授、舅舅Wilhelm His(1831-1904)为著名解剖学家。1868年春,米歇尔毕业于巴塞尔医学院后,因不感兴趣行医、自己听觉有问题、而舅舅认为“组织发育的剩余问题只能依据化学基础来解决”(Dahm,2005),米歇尔到德国图宾根接受科学训练。他在有机化学实验室工作一学期后转入Felix Hoppe-Seyler(1825-1895)实验室。Hoppe-Seyler乃时称“生理化学”(现称生物化学)的先驱,他发现血红蛋白的可逆性氧化、并命名(hemoglobin),他命名蛋白质为proteid(现称protein)。Hoppe-Seyler建议米歇尔研究淋巴细胞的化学成分,米歇尔因难以从淋巴结获足够量的纯化淋巴细胞,转而研究可大量获得的白细胞,其来源为外科诊所绷带上的脓。


米歇尔起初关注白细胞的蛋白质,他意识到蛋白质和脂肪主要位于细胞质。在研究过程中发现一种物质被酸沉淀、加碱中和后再溶于水,其特性不同于蛋白质和脂肪,他认为是新的物质,并猜测来源于细胞核,从而命名为核素(nuclein)的物质。在偶尔发现后,他重新设计实验、改进方法,用硫酸钠分离而不损坏白细胞,摸索出分离细胞核的方法,用一定比例稀盐酸去除细胞质,再反复用水和乙醚洗去附着于细胞核的脂肪,最后得到较纯的细胞核制备。用一比十万的碳酸钠处理细胞核,可以得到“物质的黄色溶液”,加乙酸或盐酸得到沉淀,碱中和后沉淀可溶(Dahm,2005)。因为这一方法获核素量不多,他探索了第二种分类核素的方法,其中用胰蛋白酶降低提取物的蛋白质含量。有足量核素后,他分析发现核素含碳、氢、氧、氮,而且含大量的磷,但不含蛋白质中常见的硫。从核素的溶解性和化学组成,米歇尔推测发现了新的物质。
1869年,米歇尔离开图宾根Hoppe-Seyler的实验室到莱比锡,在那里写好论文于当年投稿给Hoppe-Seyler主编的杂志,后者对米歇尔的结果有怀疑,直到自己和另一学生重复米歇尔的实验后,Hoppe-Seyler才同意在1871年发表米歇尔的论文(Miescher,1871),题目是“脓细胞的化学组成”(Dahm,2008),同期还刊登了Hoppe-Seyler本人及其学生Plósz的文章,两篇皆验证米歇尔的结论:Hoppe-Seyler完全肯定米歇尔的工作,并验证核素的磷含量高;Plósz验证核素只存在于鸡和蛇的有核红细胞、而不存在于牛的无核红细胞。1871年Miescher回Basel,1872年接父亲和舅舅任过的教职。在Basel,他从莱茵河三文鱼的精子提取了大量核素(Miescher,1874)。他知道核素不仅在鱼,也在蛙、牛、鸡的精子中。1872年至1874年,他提出核素中的磷都以磷酸形式存在,核素至少含有四种碱基。
以化学分析为开端的核酸研究,起初不是为了特定生物功能的分子基础。Hoppe-Seyler认为发现细胞核的物质很重要,米歇尔认为自己发现的新物质其重要性不亚于蛋白质。米歇尔发现精子中有核素后,提出“如果单个物质可以是受精的特异原因的话,那么无疑首先应该考虑的是核素”。但他又觉得不太可能是一种物质,其原因之一是核素好像不可能有很大的多样性,难以解释个体性状的多样性(Dahm,2005)。
1-2 核酸的化学分析
Hoppe-Seyler在斯特拉斯堡大学建立了德国首个生物化学系。1872年,Albrecht Kossel (1853-1927)听过Hoppe-Seyler的生理化学和病理化学课,1877年Kossel毕业于Rostock大学并考完行医执照后,加入Hoppe-Seyler的实验室,1878年开始发表有关核素的研究(Kossel,1879;Jones,1953),1883年到柏林大学工作。1885年,他从酵母的核素中发现腺嘌呤(adenine,A)。德国科学家Richard Altmann (1852-1900)获得无蛋白的核酸制备,于1889年提出核酸的名词(Altmann,1889),米歇尔认为核酸和核素相同,无需改名。Kossel用Altmann的制备进一步分析,1891年宣布发现核酸含磷酸、腺嘌呤、鸟嘌呤(guanine,G)(Kossel,1891)。1893年他和学生发现核酸含胞嘧啶(cytosine,C)、胸腺嘧啶(thymine,T)。1901年他的学生Ascoli发现尿嘧啶(uracil)(Jones,1953)。Kossel还研究了蛋白质,1884年发现细胞核中的组蛋白(histone,是二十一世纪才热门的蛋白质),1896年Kossel发现一个常见的氨基酸:组氨酸(histidine)。
|
|
|
|
|
|
|
|
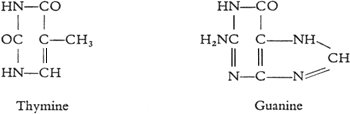
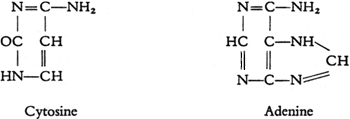

1-3 核酸的化学结构
二十世纪上半叶的核酸生物化学专家为Phoebus Levene(1869-1940)。他在俄国圣彼得堡念过军事医学院,因俄国排犹而随家人移民美国、在纽约行医,因感兴趣研究而在哥伦比亚大学注册念书,也设法获得研究训练,1896年在纽约州医院病理研究所生理化学实验室初次接触核酸。他多次到欧洲进修,曾到德国分别跟随Kossel和1902年诺贝尔化学奖得主Emil Fisher(1852-1919)。1901年John D Rockefeller(1839-1937)斥资在纽约建立与法国巴斯德研究所相媲美的洛克菲勒医学研究所。1905年Levene被第一任所长Simon Flexner聘为助理,1907年成正式研究员、并负责化学部直至1940年去世。Levene一生发表过七百多篇论文,研究过核酸、蛋白质、氨基酸、脂、碳水化合物等。
Levene发现了核酸中的核糖(ribose)、脱氧核糖(deoxyribose),提出了核酸的化学结构(现称一级结构):DNA(当初谓“胸腺核酸”)由A、G、C、T四种核苷酸(nucleotides)共价键相连而成(Levene and Jacobs,1912,1929),RNA(当初谓“酵母核酸”)由A、G、C、U四种核苷酸链接组成(Levene,1909,1917)。
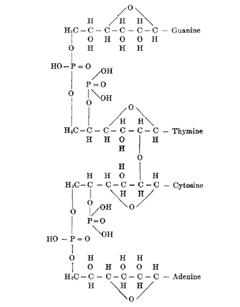


Levene最早于1909年提出核酸的“四核苷酸假说”(tetranucleotide hypothesis),起初仅强调核酸由四种核苷酸组成,反驳其他人提出二核苷酸、三核苷酸假说(Levene,1919,1920a,1920b)。在他1912年提出DNA的结构时显示了四种核苷酸,但未提四种核苷酸的相对含量。后来四核苷酸假说被推广为DNA链中各种核苷酸的含量相同(A:T:G:C=1:1:1:1),是单调重复的分子、不太可能为信息的载体。
2 核酸与染色质
2-1 核酸的亚细胞定位
1914年,德国的Robert Feulgen(1884-1955)发现DNA在溶液中通过盐酸(暴露出DNA的醛基)和Schiff试剂(品红亚硫酸,可与醛基反应)两步可显紫红色,RNA不能显色,后称Feulgen反应(Kasten,2003)。
1923年,Feulgen将这一反应引入组织化学:直接在生物的组织切片上进行反应,以此确定DNA在组织或细胞的存在部位。
1924年他和技术员Heinrich Rossenbeck以此方法检测多种动植物组织、细胞后证明DNA存在于细胞核,不仅动物细胞核、而且植物细胞核(Feulgen and Rossenbeck,1924)。
Feulgen也改变了前人误以为DNA(胸腺核酸)存在于动物、RNA(“酵母核酸”)存在于酵母和植物,而从酵母中提取DNA还要到1948年(Chargaff and Zamenhof, 1948)。
2-2 核酸与染色质
瑞典卡罗琳斯卡医学院的 (1910-1997)发现核酸对260nm紫外线有最佳吸收峰(Caspersson,1932,1936),Morgan以前的学生Jack Schultz 与Caspersson用紫外检测将DNA定位于细胞核(Schultz and Caspersson,1940),他们还观察到果蝇唾液腺多线染色体条带变化后核酸含量变化、果蝇卵母细胞染色体数量变化可以改变核酸含量(Caspersson and Schultz,1938)。
(1910-1997)发现核酸对260nm紫外线有最佳吸收峰(Caspersson,1932,1936),Morgan以前的学生Jack Schultz 与Caspersson用紫外检测将DNA定位于细胞核(Schultz and Caspersson,1940),他们还观察到果蝇唾液腺多线染色体条带变化后核酸含量变化、果蝇卵母细胞染色体数量变化可以改变核酸含量(Caspersson and Schultz,1938)。
Caspersson与同事Einar Hammarsten(1889-1958)合作分析染色体的核酸和蛋白质组分,用蛋白酶消化蛋白质后得到高纯度的核酸(Caspersson,Hammarsten,Hammarsten,1935)。他们观察到果蝇多线型染色体条带与核酸的关系非常逼近核酸与遗传的关系。
Hammarsten和Caspersson发现DNA不是短链、而是长链,分子量很大(50万到100万),所含嘌呤环和嘧啶环的平面与链的长轴垂直(Signer, Caspersson, Hammarsten,1938)。英国Leeds大学纺织物理实验室的William Astbury 通过X线衍射分析发现垂直于长轴的两个核苷酸之间距离为3.34Å(Astbury and Bell,1938),Astbury认为核苷酸间距与蛋白质中氨基酸间距很接近可能非巧合、而说明核酸链与蛋白链之间有关系。
1942年,比利时的Jean Brachet(1909-1988)用染色方法证明DNA在细胞核的染色体上,RNA在动物细胞质与核仁中(Brachet,1942;Thomas,1992)。他用吡罗红(pyronin)和甲基绿(methyl green)混合染料,甲基绿染DNA显绿色,吡罗红染RNA显红色,两者都染显蓝色。当时俄国移民美国在洛克菲勒医学研究所工作的生物化学家Moses Kunitz(1887-1978)利用RNA酶(RNase,降解RNA)对热不敏感的特性,从牛胰腺提取到高纯度的RNase(Kunitz,1940)。Brachet用RNase处理组织切片,可以去除RNA的染色、只显DNA染色。他发现染色体含DNA(可看到有条带的昆虫巨大染色体上DNA染色也呈条带,有些条带还含RNA),细胞质含RNA、且其含量与蛋白质含量有相关性。
至1940年代初期,确切知道DNA存在于细胞核的染色体上。不过,染色体上即能检测到DNA、也能检测到染色体上的RNA和蛋白质,并不能仅仅由亚细胞定位确定DNA是遗传物质。
3 蛋白质与核酸
染色体既含核酸、也含蛋白质,携带遗传信息的分子究竟是什么?
二十世纪上半叶,一般认为蛋白质很重要。1934年,英国的J D Bernal(1901-1971)和Dorothy Hodgkin(1910-1994)第一次获得蛋白质(胃蛋白酶)的晶体结构(Bernal and Crowfoot, 1934),显示蛋白质的结构复杂性。此前已知蛋白质生化特性和功能多种多样,人们易信蛋白质可以携带丰富的信息。
酶为生物催化剂的概念于十九世纪提出、到二十世纪初还争论酶是蛋白质还是其他分子。因研究叶绿体而获1915年诺贝尔化学奖的德国犹太科学家Richard Willstatter(1872-1942)认为酶是蛋白质制备中的其他污染物质。其他科学家的工作,特别是1926年美国Cornell大学的James Sumner(1887-1955)和1930年洛克菲勒医学研究所的John Northrop(1891-1987)分别获得结晶纯的尿素酶和胃蛋白酶,证明酶的分子本质是蛋白质,获1946年诺贝尔化学奖。在这样的背景下,已知染色体有蛋白质和核酸时,很多人怕再次犯低估蛋白质重要性的错误(Judson,1979)。
研究核酸的生物化学专家Levene认为核酸单调重复,最初以为就是四种核苷酸,曾以为核苷酸形成的链不长,但即使在Signer, Caspersson, Hammarsten(1938)确定DNA分子量很大、是很长的链以后,仍未及时改变核酸像淀粉一样不太可能携带信息的错误观念。
用染色确定DNA存在于细胞核中的Caspersson、Hammarsten、Brachet皆未提出核酸是遗传的物质基础。Caspersson认为染色体中的蛋白质可能是遗传物质(Caspersson,1936)。
1941年Schultz认为按遗传学的预计基因应该是线性、有特异性、存在于染色体上、能自我复制、同时能影响细胞的合成代谢,而核酸与蛋白的复合体(nucleoprotein)符合这些条件、应该是基因的物质基础(Schultz,1941)。他在分别讨论染色体的核酸和蛋白质时,认为蛋白质确实有特异性,而核酸是否有特异性尚不清楚:虽然一般以为核酸单调,他指出当时分析过结构的核酸只有来源于胸腺的,不能排除不同细胞的核酸有特异性的可能性。1943年,他指出病毒肯定有基因,从病毒、细菌到高等动植物都有核酸和蛋白质复合体,而它们都能复制,所以细菌是否有细胞核并不重要,有核酸和蛋白质复合体为基础的基因。他依据已知三种含氨基酸不同的烟草镶嵌病毒(TMV)所含核酸在当时检测显示很恒定,提出应该是蛋白质给予病毒特异性、而核酸不能(Schultz,1943),所以最接近提出核酸是遗传基础的Schultz因当时检测手段的限制退而认为蛋白质是遗传物质。
4 肺炎球菌的分型和分类
4-1 肺炎球菌的分型
十九世纪下半叶,法国的巴斯德(Louis Pasteur,1822-1895)和德国的科霍(1843-1910)创立了现代微生物学:继承前人积累确立了细菌致病论(germ theory)、发现重要微生物、发明预防传染病的疫苗、培养微生物学家。
细菌是单细胞生物,但无典型的细胞核,所以称为原核生物(prokaryotes)。细菌细胞质外有细胞膜(plasma membrane),膜外有细胞壁(cell wall),有些细菌还有荚膜(capsule)。
细菌性肺炎长期困扰人类、在抗生素应用以前常导致死亡,二十世纪初美国每年因肺炎去世逾5万人。即使现在全世界每年也有上亿肺炎患者,肺炎是疾病导致儿童死亡的最大原因,每年近百万5岁以下儿童死于肺炎(Henriques-Normark and Tuomanen, 2013)。1918年,著名内科医生William Osler(1849-1919)称肺炎为“人类死亡的罪魁祸首”(captain of the men of death)。肺炎可以通过空气传染,其流行为国家所关注。
1884年,德国的犹太医生Albert Frankel(1864-1938)发现肺炎球菌(Streptococcus pneumoniae)。
肺炎球菌可以分型,其中主要方法依据抗原性(antigenicity):二十世纪初可以针对肺炎球菌制备含不同抗体(antibodies)的抗血清(antiserum),含不同抗原的肺炎球菌与不同的抗体反应。德国细菌学家Friedrich Neufeld(1869-1945)提出这些抗体既可抑制特定类型的肺炎,又可用来将肺炎球菌分成I、II、III型(Neufeld and Levinthal, 1909,1912)。
美国洛克菲勒研究所附属医院的Alphonse Dochez(1882-1964)发现肺炎球菌还有IV型,且IV型中还有多种(Dochezand Gillespie, 1913)。1913年Oswald Avery(1877-1955)加入洛克菲勒,1917年他与Dochez等发现IV型更多种类(Avery et al., 1917)。1922年,英国卫生部病理实验室的Fred Griffith(1879-1941)发现IV型中至少有12种不同的类型(Griffith,1922)。I型和II型常致病强,IV型致病弱、存在于有些正常人的口中,IV型内不同株的差异较大、有些也有致病性。一般来说,一种肺炎球菌只含这四型的一种抗原。
不同抗原型的肺炎球菌因何不同?1917年,Dochez和Avery发现肺炎球菌的抗原不是肺炎球菌细胞破裂后从细胞内掉到细胞外的物质,而是细菌表面的物质,可溶于培养液和体液(Dochez and Avery,1917)。Dochez一战入伍离开洛克菲勒,Avery实验室1923年至1934年一系列实验阐明肺炎球菌分型的抗原不是蛋白质、而是多糖(polysaccharide)(Heidelberger and Avery, 1923;Avery and Goebel, 1933; Goebel, Avery and Babers, 1934),这些抗原为细菌细胞荚膜的完整性所需,也就对肺炎球菌的致病性重要,Avery等的研究是免疫化学的重要工作。
4-2 肺炎球菌类的转换
细菌的类型能否变化?1887年,细胞免疫学创始人、俄国的Ilya Metchnikoff(1845-1916)发现炭疽杆菌在抗血清中培养后致病性降低。1915年南非的AR Friel发现肺炎球菌在抗血清培养后降低致病性并改变抗原性。1916年,Avery实验室的研究生Laura Stryker发现肺炎球菌的致病性经抗血清培养后下降,而进入动物体内可以恢复(Stryker,1916)。1921年,伦敦Lister研究所的Joseph Arkwright(1864-1944)总结他研究的几种肠道细菌,它们有两种形态,根据表面光滑程度被Arkwright称为S类和R类。
1922年, Griffith向英国卫生部报告1920年至1922年150例大叶性肺炎病人中肺炎球菌分型的情况(Griffith,1922)。1923年,Griffith向卫生部报告,致病的(virulent)表面光滑(S类),不致病的(avirulent)表面粗糙(R类),它们有不同抗原性(可以分别有抗S的抗血清和抗R的抗血清)。在体外培养肺炎球菌时如有抗血清,肺炎球菌的致病性和形态会改变,从S类变成R类,还观察到R类变成S类(Griffith, 1923)。
1925年,洛克菲勒的Hobart Reimann验证在血清或胆盐、甚至一般培养基中培养I型S类的肺炎球菌后,致病的S类细菌有些变成R类,失去致病性、改变形态和抗原性,但在他的实验中R类无论在体外还是动物体内都不能变成S类(Reimann,1925)。霍普金斯大学的Harold Amoss也验证了Griffith的结果:致病类的I型肺炎球菌,可以在体外培养变成不致病类;但Amoss未观察到不致病类回复为致病类(Amoss,1925)。因为Reimann(1925)、Amoss(1925)用了分离的单菌落做实验,较Stryker(1916)和Griffith(1923)更有说服力,证明单个S类的菌变成R菌、而不是最初S类菌落中混有R菌个体。Reimann还发现有些型的肺炎球菌也可以在动物(如兔、豚鼠、马,但非狗)体内从S类变成R类(Reimann,1927)。
5 肺炎球菌型间转化实验
1928年,Griffith在《卫生学杂志》上发表了细菌转化的论文(Griffith, 1928)。首先,他分析了1920年至1927年的流行病资料,发现如果将病例分成三个时间段(1920至1922、1922至1924、1924至1927),那么随着时间推移,感染I型和III型的人数无规则性变化,但II型的减少(从32.6%变为7.4%)、IV型的增多(30.0%变为53.7%)。
他注意到1922年的病例中单个病人可以同时携带不止一型的肺炎球菌。这是因为病人多次感染,还是只有一次感染一型的肺炎球菌、其后细菌在病人体内从一型变成另一型?1922年到1927年病人群体中观察到的趋势支持变型的可能,但仅从相关性进行推测,缺乏直接证据。
Griffith从1922年一位含多型肺炎球菌之病人的痰中获得I型肺炎球菌,接种老鼠可以致病,从老鼠中获得肺炎球菌再接种下一群老鼠,这样系列传代(serial passage),发现后来可以出现IV型的肺炎球菌,从而显示I型可以变成IV型。他再从多位病人获得I型肺炎球菌、在老鼠接种验证可以变成IV和III型,从多位病人获得II型肺炎球菌、在老鼠可以变成III型。


这时,他用典型的英国双重否定句:综合考虑可能性来看,型间转换假说的可能性似乎不比多重感染更不可能(on a balance of probabilities, interchangeability of type seems a no more unlikely hypothesis than multiple infection)。
Griffith认为已有证据还不足以区分型变与多重感染两种假说,需要继续实验。
他发现,一般来说无论肺炎球菌依据抗原分型是哪种(I、II、III或IV),都有致病的S类和不致病R类。无论哪种抗原分型,其致病的S类如果在老鼠传代接种,都可变成不致病的R类。也有两种方式可在体外培养让S类变成R类:一种是将某型的S类细菌(如I型的S类)在相应的抗血清中培养(如含I型抗血清),其后可获同型的非致病菌(如I型的R类);另一方式是不加抗血清,在固体培养基中培养,传多代后也能从S类获R类。不过每次的菌株变化的情况不同、传代后的菌株稳定性也不同,有些变成R类后可以很快回复为S类,有些变成R类后很稳定、难以回复为S类。回复变化(reversion)只能在同一抗原分型之内,如:I型的R类变成I型的S类,不出现I型的R类变成II型的S类。保存在冰箱但未被允许生长分裂的肺炎球菌不出现R类和S类之间的变化。
Griffith将II型的S类菌加热到100°C,灭活其致病性,而II型的R类菌已知无致病性,但II型的S类加热后、与II型的R类菌同时注射到老鼠时,可以致病、而且从患病死去的老鼠可获II型的S类菌。如果II型的R类与加热100°C处理的I型S类菌同时注射老鼠,结果无致病性,似乎同型的S类加热灭活后可以转化同型的R类。但Griffith用加热的I型S类菌与II型的R类菌共同注射老鼠时,可以致病,不过加热时的温度需要注意,60°C可以、而100°C会灭活I型S类菌的转化活性。所以,在一定条件下不同抗原型的肺炎球菌,在加热失去致病性后,仍可将另一抗原型中无致病性的R类细菌转化为可致病的S类细菌。如果两类型都加热了,同时注射无致病性。Griffith用60°C和100°C加热处理过其他型的S类肺炎球菌,然后检测其结果。他发现多数型的S类都能转变其他型的R类(如加热的I型S类变II型R类为I型S类、加热的II型S类变I型R类为II型S类、加热的III型S类变I型R类为III型S类、加热的I型S类变IV型的R类为I型S类),少数转化不行(如IV型的S不能变I型的R为IV型的S,但能促进I型的R回复为I型的S)。一型的R类加热后不能转化另一型的R类、也不能促进另一型的R类回复为S类。
Griffith提出,S类的菌含S物质,S物质是蛋白质,它帮助制造细菌荚膜有抗原性的多糖。他当时认为,I型的肺炎球菌同时有I型和II型的S抗原,只是I型的细菌含I型的S抗原多于II型的S抗原,而R类的I型细菌其S抗原大多数变成了R结构,在加热后的I型S类细菌提供S物质的情况下,II型R细菌获得I型的S抗原、获得致病性,也可因加热的I型S类菌含少量II型的S抗原,后者转II型R类菌荚膜导致II型R类菌回复为II型S类菌。
Griffith的解释是从细菌抗原和致病性的角度,没有考虑遗传突变、回复突变和基因转导,但他一步一步从病例观察到提出和证明细菌转化,做出了关键的贡献。可惜,1941年Griffith和同事一道逝于德国飞机轰炸。
6 Avery实验室的主线:诊断、治疗和预防
Avery实验室主要研究目的是在理解肺炎球菌的基础上,帮助诊断、治疗和预防细菌性肺炎,这些目的多数达到了、但时间不同,有些为几十年后他的学生的学生做到。
1877年出生于加拿大的Avery,十岁移民美国纽约,学过医但对研究更感兴趣,特别对危害人类的传染病(如肺结核)。1913年,洛克菲勒医学研究所附属医院的院长Rufus Cole(1872-1966)看重Avery在传染病方面的研究论文,请他加入洛克菲勒,Avery从此精力集中于肺炎球菌。
在理论方面,1916年Dochez和Avery提出“抗生长免疫”的概念(antiblastic immunity),认为抗体抑制肺炎球菌细胞内的酶活性,从而抑制代谢(Dochez and Avery,1916)。此理论当时就不为洛克菲勒内部其他学者(包括院长)所认同,一般认为抗血清导致细胞凝集后继发细菌生长减慢、代谢降低。Avery这次错误使他以后更为谨慎。
在诊断方面,Avery实验室从1923至1934年阐明细胞荚膜多糖抗原后,用细胞荚膜多糖抗原分型仍为今天追踪肺炎球菌的方法,到2013年知道有93型(Henriques-Normark and Tuomanen,2013),所以Avery的研究对诊断有帮助。
在治疗方面,Avery在发现荚膜多糖对致病的重要性以后,寻找降解多糖的方法,希望通过降解多糖、破坏荚膜来治疗肺炎。他于1927年在一次午餐时偶遇Rutgers大学毕业的研究生René Dubos(1901-1982),Dubos来自有土壤细菌专长的实验室、原导师为Selman Waksman(1888-1973)。Avery与Dubos一拍即合,觉得细菌在土壤里应该分解,所以在土壤里有降解多糖的分子,几年后他们确实找到了降解多糖的酶(Avery and Dubos,1931),而且可治疗动物的肺炎。不过,用蛋白质性质的酶作为治疗药物,不如小分子的抗生素,所以未得到广泛应用。
在预防方面,1914年Wright等就曾为南非矿工制造疫苗以预防肺炎(Wright et al., 1914)。真正得到公认有效的肺炎疫苗是Avery的学生Colin MacLeod有独立实验室后于1945年制造的针对多糖的治疗性抗血清(MacLeod et al., 1945)。不巧正好抗生素被迅速推广,而青霉素成为治疗肺炎的标准药物,很多人以为无需疫苗预防,所以多糖的抗血清当时未得应用。但是,1964年MacLeod的学生Robert Austrian(1916-2007)发现细菌性肺炎仍有很高致死率(Austrian and Gold,1964),重新提出研发和使用疫苗的必要性。Austrian再度依据多糖抗原发明了多效价疫苗,成功地预防儿童肺炎(Austrian et al., 1976),这一方法也是迄今疫苗的主流,所以在预防方面Avery本人播种而结果于他去世二十多年后他的学生的学生。
Avery实验室有心栽花是诊断、治疗、预防,有些开了、有些没开、有些很晚开了。
7 Avery实验室的支流:转化和遗传
Avery实验室做转化实验初为无心插柳,最后成荫、成林…到今天是漫山遍野的万紫千红。
细菌转化实验是Avery实验室的支流。发现了现象最初不知其意义多大,断断续续地追踪,但最后意义远超出细菌性肺炎一种传染病,而是整个生物学的革命,也改观了传染病和非传染病的诊断、治疗和预防。
1916年,Avery实验室的Stryker就知道致病性和非致病性肺炎球菌的变化(Skryker,1916)。
1923年Griffith第一次初步报道致病性和非致病性的肺炎球菌是S和R类后,Avery实验室的Reimann于1925和1927年重复了Griffith的研究结果(Reimann,1925,1927)。
1928年Griffith经典的转化实验发表前,德国柏林细菌研究所的Neufeld访问Griffith实验室得知其结果并进行了重复(Downie,1972),同年Neufeld发表验证文章(Neufeld and Levinthal, 1928)。1927年10月离开纽约到洛克菲勒基金会支持的北平协和医学院内科任副教授的Reimann,很快在北京继续肺炎研究,1928年10月提交的文章于1929年发表(Reimann,1929):他验证了Griffith的结果,而且用Reimann于1925年发现的不能自身在体外或体内变成S类的I型R类细菌,用Griffith的方法可以被转化为S类细菌,其抗原型取决于用来做转化的加热处理过的S类细菌的抗原型(I、II、III)。
Griffith文章发表后,Avery对其有怀疑。那几年他自己的注意力集中于寻找降解多糖的分子。
Avery实验室从英属加拿大区来的博士后Henry Dawson (1896-1945)相信英国人Griffith的结果,并在体内重复验证(Dawson,1928,1930),但他试图在体外做转化实验未成功(Dawson,1930)。Dawson搬到哥伦比亚大学有独立实验室后,与协和医学院赴美国进修的谢和平(Richard Ho-Ping Sia)在体外细胞培养的条件下进行转化实验,也就是看加热后的一型S类的细菌能在体外共培养时转化另一型R类细菌(Dawson and Sia,1931;Sia and Dawson,1931)。但Sia与Dawson用肺炎球菌的提取物未能做成转化(Sia and Dawson,1931)。已知用来分型的抗原为多糖,而R类有多糖的缺陷(才造成细胞荚膜缺陷),但将S类的多糖加到R类的细菌却不能将R类变成S类,所以转化物质不是多糖(Sia and Dawson,1931)。
1932年,Avery实验室的James Lionel Alloway发现用I(或III)型的S类肺炎球菌的提取物可将II型R类肺炎球菌转化为I(或III)型S类肺炎球菌(Alloway,1932),60°C到90°C加热处理不能灭活提取物的转化活性。Alloway也指出用S的多糖不能转化R(Alloway,1932)。1933年,Alloway继续摸索提取的条件(包括脱氧胆酸钠sodium deoxycholate萃取能得到更多活性物质、木炭可以吸附、丙酮或乙醇可沉淀),观察转化物质的化学性质,发现提取物无抗原性,所以肯定不是多糖抗原本身(Alloway,1933)。
8 转化因子(Transforming Principle)
Dawson和Alloway离开Avery实验室后,Edward S Rogers从1932到1934继续他们的研究。1934年,加拿大的Colin MacLeod(1909-1972)到美国加入Avery实验室,研究继续Dawson和Alloway的课题,从S类细菌的提取物中获得转化R类细菌活性物质。
起初R类细菌本身不稳定(容易变成同型的S类),而S类提取物的活性也不稳定。MacLeod花了三年时间探索条件提高稳定性。1931年谢和平与Dawson发现稳定性低的R类不仅易自身回复为同型的S类(I型R类回复为I型S类)、且易被异型的S类(如II型的S)转化(为II型的S类)(Sia and Dawson,1931)。不稳定的R类不很合适用来检验S的转化活性。MacLeod用II型S类一株菌传36代后,获得R36A株菌,非常稳定、不易自身回复为同型S类,但却仍可被加热的异型S转化为异型S类。MacLeod也探索了改进S类提取的方法。但到1937年,他在转化实验方面的进展很慢。他还做了其他研究、发表了文章。1940年MacLeod和Avery两人都做提取转化因子的实验,改进了方法,如用Sevag (1934)的氯仿法去除蛋白质、先加热(后来知道加热灭活细菌的DNA酶防止DNA降解)再提取,他们于1941年1月28日第一次记录转化活性的粗提物中有DNA(和RNA)。1941年7月,MacLeod离开Avery实验室到纽约大学任微生物系主任。
Avery患Graves病引起甲状腺亢进,1934年做甲状腺切除术后短时休息,多年未完全康复。1935年至1941年Avery无论文(实验室有,但他未做作者),1941年有三篇论文(皆非转化方面),1942和1943年无论文。1943年Avery到了强制性退休年龄,但二战期间美国怕出现一战时传染病导致军队大量减员的情况而请微生物学家为国家工作,洛克菲勒医学研究所也给Avery荣誉研究员,但实际继续主持实验室。
一位Avery的学生称自己在1936年的笔记上记载Avery当时说过转移因子不是碳水化合物、也不像蛋白质,而可能是核酸(Hotchkiss,1965)。
1941年9月,Maclyn McCarty(1911-2005)加入Avery实验室。McCarty在Stanford念本科、Hopkins念医学院期间都参加过生物化学研究,提取分离过蛋白质。Avery给他的课题是提取转化因子,McCarty首先用多糖的降解酶证明转化因子本身不是多糖,并在生化上有很多改进,到1942年夏天多方面结果指向转移因子是DNA,所以专门提取肺炎球菌的DNA,大规模提取时每次要两、三百升的细菌,1943年做完实验,暑期Avery开始写文章、11月投稿。
1944年Avery、MacLeod、McCarty共同发表论文(Avery, MacLeod and McCarty,1944),一般认为与Watson和Crick(1953)并列为二十世纪生物学最重要的文章。
Avery-MacLeod-McCarty开篇高屋建瓴,完全不同于前面一系列限于肺炎球菌分型的文章,而是将问题提高到遗传的分子机理层面:首先指出很多生物学家试图用化学的方式在高等生物诱导特异、可预见、能遗传的变化。其次认为微生物中可以实验诱导、并重复发生、可遗传的特殊变化最好的例子是肺炎球菌型的转化实验。其后他们简介Griffith的实验结果,以及Dawson、Alloway等的进一步工作。Avery-MacLeod-McCarty的研究目的是找有转化活性的因子(principle),并初步确定其化学性质、化学分子的类型。
达到宏大的目标需要能解决问题的具体方法。用来获得提取物的是III型S类菌株(A66),与被转化的II型R类有很大的形态和大小差别。转化实验用的培养液,MacLeod用炭可去除不稳定性。转化实验仍需血清,以前谢与Dawson认为血清提供抗R的抗体,Avery等认为与抗体无关,但仍需血清的某种其他成分,因血清含灭活转化活性的酶,需将血清加热到60°C,灭活此酶。用于检测转化活力的细菌,需不易自身转变、而同时对转化活性有反应,这就是MacLeod用II型S传36代后获得的R36A菌株。他们注意到即使是R36A还可衍生不同的菌株,有些可被转化、有些不能,而R菌破损释放的酶能降解S菌的转化活性,所以需选生长良好的R以免检测的不稳定。他们采用Sevag(1934)发明的方法用氯仿去除蛋白质、接着用乙醇多次沉淀,用Avery实验室发现的降解多糖的酶去除多糖,再逐滴用乙醇达到临界浓度沉淀活性物质,得到粘性的纤维,用玻璃棒搅出来再溶解。乙醇沉淀DNA的方法是洛克菲勒生化学家Alfred Mirsky告诉McCarty(Mirsky and Pollister,1946;Darnell,2011)。
提取的活性物质在生理盐水中很稳定可保存数月、但在水中不稳定,65°C一小时不能灭活,用两种检测蛋白质的方法(双缩脲和Millon检测)结果显隐性,用DNA的检测方法(Dische氏二苯胺反应)显强阳性,RNA检测方法(Bial氏地衣酚反应)显弱阳性,但他们用同样浓度的提取的胸腺DNA看到用地衣酚反应也显弱阳性。反复用乙醇和乙醚提取而不降低活性,推测活性分子不是脂肪。
他们对四批提取的转化物质进行元素含量的检测,含C约34%、H约3.8%、N约15%、P约9%,都较接近纯化的胸腺DNA中元素的含量(34%、3.2%、15%、9%)。N/P比例为1.58到1.75,也接近DNA预计值1.69。
他们从洛克菲勒的酶专家Northrop和Kunitz获结晶纯的胰蛋白酶、糜蛋白酶和RNA酶,它们都不能灭活转化因子,说明转化活性非RNA、也非这些蛋白酶能降解的蛋白质。
他们从几种动物组织制备粗提物,a狗小肠粘膜、b兔骨磷酸酶、c猪肾磷酸酶、d肺炎球菌自溶物、e狗和兔血清。在酶活性方面,这些粗提物除d外都有磷酸酶活性、除c外都有甘油三丁酸酯酶活性、除b和c外都有DNA酶活性(检测DNA酶活性的DNA由洛克菲勒的Mirsky提供),而只有a、d和e能够灭活转化物质,比较磷酸酶、酯酶和DNA酶活性的相关性倾向于支持DNA是转化因子。
他们用不同温度和时间处理狗和兔血清,观察到热灭活DNA酶活性负相关于这些制备对转化因子的灭活。而温度灭活血清中甘油三丁酸酯酶的活性与转化活性无关。
他们试了一些化合物能否抑制灭活转化因子的酶,发现氟化钠有显著作用,它可以抑制所有已知来源(如肺炎球菌、狗小肠粘膜、胰腺、血清)的DNA酶的活性,也抑制这些来源DNA酶制备物对转化因子的灭活,进一步支持DNA与转移因子的相关性。
转化因子本身对III型抗血清无反应,不是III型的抗原本身,也就不是荚膜多糖。
他们请生物物理学家分析,发现在超速离心中,转化物质显示均质性和非对称性,分子量约50万。电泳行为也显示类似核酸的一种分子。UV光谱显示吸收峰在260nm,也与核酸一致。
转化因子活性很强:2.25毫升溶液含0.003微克仍有活性,六亿分之一的比浓度。
在讨论中,他们说从III型肺炎球菌分离了DNA组分可以将II型衍生的无荚膜R类菌转化为完整荚膜的III型菌。肺炎球菌有RNA是1938年洛克菲勒的`科学家所发现(Thompson and Dubos,1938),而1944年前尚无人报道肺炎球菌的DNA。他们强调转化因子的化学性质不同于其导致产生的荚膜多糖。他们指出“本文所呈现的资料强烈提示核酸,至少是脱氧类型的,具有转化因子选择性作用所证明的不同特异性”。他们也知道细菌一旦被转化后,获得的性状可传代,转化带来的变化“可预计、有型的特异性、可遗传”。
对转化的机理,Avery-MacLeod-McCarty讨论了三种,其中第二种是遗传学家Dobzhansky(1941)提到的转化可能是遗传突变(如果转化被描述为遗传突变—很难避免如此描述—我们就在面对特别处理导致特异突变的真实例子),Avery-MacLeod-McCarty认为这是比拟转化因子是基因。
他们承认还可能有其他物质吸附在核酸上起作用,但如果确实能证明就是核酸起转化作用,那么核酸应该具有尚未确定化学基础的生物学特异性。
全文的结论:出示的证据支持脱氧类的核酸是III型肺炎球菌转化因子的根本单位(the evidence presented supports the belief that a nucleic acid of the desoxyribose type is the fundamental unit of thetransforming principle of Pneumococcus Type III)。



1945年,英国神经生理学家、1936年诺贝尔奖得主Henry Dale(1875-1968)在给Avery发英国皇家学会的Copley奖时,提到Avery发现了基因(Dale,1946),而诺贝尔奖委员会受其研究核酸的成员Hammarsten影响直到1952年以前都不认同DNA是遗传的物质基础(Reichard,2002)。
终生未娶的Avery于1947年离开纽约去田纳西州,在任Vanderbilt大学微生物教授的弟弟Roy Avery家附近过退休生活,1955年去世。
9 对新概念的反应
当时的科学家如何理解1944年Avery-MacLeod-McCarty的文章?知道Avery-MacLeod-McCarty等工作的科学家面临三种很重要的选择:不理不睬、检测真伪、走下一步。
9-1 很难排除DNA之外完全无蛋白质
十九世纪核酸的生物化学权威是Miescher和Kossel,二十世纪1940年以前核酸的生化权威是洛克菲勒的Levene,而1940和1950年代核酸生化的权威为Alfred Mirsky(1900-1974)。Mirsky于1927年加入洛克菲勒医学研究所前期研究蛋白质(特别是血红蛋白),1940年代后主要研究细胞核的核酸和蛋白质。二十世纪上半叶两位核酸生化权威Levine和Mirsky皆低估核酸的重要性。
Mirsky和哥伦比亚大学的Arthus Pollister主要依赖不同浓度的NaCl,从精子、肝、胰、肾等将细胞核中核酸和蛋白质的复合体(时称“核蛋白”nucleoprotein)与细胞质分开,获纤维状物质(Mirsky and Pollister,1942)。他们还能分开复合体的核酸和蛋白。Mirsky改进方法后不仅可从动植物等真核细胞获得细胞核的核蛋白,且可从显微镜看不到细胞核及染色体的原核生物(细菌)中提取核蛋白,他们提出所有细胞都有DNA和组蛋白,在组蛋白以外染色体还含其他蛋白质(Mirsky and Pollister, 1946)。他们用来提取核酸和蛋白质的细菌正是Avery给他们的III型肺炎球菌,并提到Avery-MacLeod-McCarty提取转化因子的方法部分参考了Mirsky和Pollister方法(Mirsky and Pollister, 1946),与McCarty后来的回顾一致(Darnell, 2011)。Mirsky和Pollister(1946)指出核蛋白的纤维状是因多聚体的DNA,而非1937年诺贝尔奖得主 (1893-1986)等误认为与肌球蛋白相似。他们也用提取的核蛋白(当时他们还曾称为chromosin)验证了Avery-MacLeod-McCarty的转化实验,但他们指出很难获得完全不含蛋白质的纯DNA,而当时的方法无法检测出低于2%的蛋白质,所以不能断定转化活性是DNA、还是蛋白质,他们认为还需一步一步去除蛋白质、同时检测转化活性(Mirsky and Pollister, 1946)。他们曾认为可以用氯仿-辛醇反复6、7次去除蛋白质、再用乙醇沉淀后获纯化的DNA(Mirsky and Pollister, 1946),但他们未进一步用自己提出的方法来验证转化因子是核酸还是蛋白质。
(1893-1986)等误认为与肌球蛋白相似。他们也用提取的核蛋白(当时他们还曾称为chromosin)验证了Avery-MacLeod-McCarty的转化实验,但他们指出很难获得完全不含蛋白质的纯DNA,而当时的方法无法检测出低于2%的蛋白质,所以不能断定转化活性是DNA、还是蛋白质,他们认为还需一步一步去除蛋白质、同时检测转化活性(Mirsky and Pollister, 1946)。他们曾认为可以用氯仿-辛醇反复6、7次去除蛋白质、再用乙醇沉淀后获纯化的DNA(Mirsky and Pollister, 1946),但他们未进一步用自己提出的方法来验证转化因子是核酸还是蛋白质。
1947年,Mirsky在提取染色体后确定DNA占其质量的37%和蛋白占59%(Mirsky and Ris, 1947)。他们继而发现同一生物的不同体细胞中DNA含量相同,而体细胞DNA含量是精子DNA的两倍,因为体细胞是二倍体有两套染色体、而精子是单倍体,所以DNA含量与染色体数量相关(Mirsky and Ris, 1949, 1951)。这虽进一步支持DNA是基因,但Mirsky至1951年仍不信仅由DNA而无蛋白质携带遗传信息(Mirsky,1951)。
9-2 纯DNA的转化活性
Avery-MacLeod-McCarty(1944)认为转化因子是DNA非蛋白质的证据之一是DNA酶可以降解转化因子,而蛋白酶、RNA酶不能。但是,当时用的DNA酶并非纯化的DNA酶,而是很粗糙的制备,含很多其他的酶,Avery-MacLeod-McCarty只比较了这些制备物含磷酸酶、酯酶和DNA酶的相对分布,所以他们也承认有关转化因子的酶学证据是间接的(McCarty and Avery, 1946a)。实际上,DNA酶不过是1940年才由美国国立健康研究院癌症研究所的Jesse Greenstein和Wendell Jenrette所发现、1943年命名,要到1948年洛克菲勒的Kunitz才能获结晶纯的DNA酶(Kunitz,1948)。1944年Avery-MacLeod-McCarty文章发表时,全世界谁也没有纯化的DNA酶。
为解决这一问题,1946年McCarty从牛胰获相当纯化DNA酶、活性很高,不含RNA酶、酯酶、磷酸酶、但含很低的蛋白水解酶活性(McCarty, 1946a)。镁离子可激活、柠檬酸可抑制DNA酶,特异的抗血清也可抑制DNA酶(McCarty, 1946a)。用经过一定纯化制备的DNA酶,他们再检验转化因子是否DNA(McCarty and Avery, 1946a)。这次,他们也观察了相关性。镁离子激活DNA酶活性,但镁的作用可以被柠檬酸抑制,而锰离子也激活DNA酶活性,但锰的作用不被柠檬酸抑制。而这些作用都与柠檬酸能否抑制镁离子和锰离子对转化活性的作用相关。对DNA酶制备物还含微量蛋白水解酶活性的问题,他们用稀释的方法予以排除。制备物在含蛋白质每毫升0.2毫克以上时可以检测到蛋白水解酶的活性,但他们稀释到蛋白质浓度低于每毫升0.01微克、直至每毫升含0.0025(甚至0.00125)微克时,还有DNA酶的活性和降解转化活性的作用,而这时无蛋白水解酶活性(需要浓度提高十万倍才有),这样的结果应该很难否定转化因子是DNA的可能性。
发现柠檬酸对DNA酶的抑制作用后,McCarty和Avery再重新提取III型S类菌的转化因子,在提取过程中加柠檬酸,提高了转化因子产量五倍(McCarty and Avery, 1946b)。他们从II型和VI型肺炎球菌也找到了转化因子,对II型转化因子的分析也支持是DNA。
1946年Rollin Hotchkiss(1911-2004)加入转化因子研究,至1949年可以做到提取的转化物质中蛋白质含量低于0.02%却仍有转化活性,而且在纯化过程中虽然蛋白质含量越来越低、但转化活性不降低,如果转化因子还是蛋白质,那就非常不同于其他蛋白质(Hotchkiss, 1952)。
9-3 核酸的特异性和信息承载的可能性
Levene的“四核苷酸假说”导致误解:核酸单调无信息含量。
核酸生化权威如此,而在二战前后都研究核酸生物物理的英国科学家William Astbury(1898-1961)也认同四核苷酸假说(Astbury,1947)。
哥伦比亚大学的旅美奥地利犹太生化学家Erwin Chargaff(1905-2002)受Avery-MacLeod-McCarty(1944)结果的激动,从脂类研究转向核酸研究,用生化方法检测不同组织、细胞来源的DNA是否含等量的四种核苷酸。他实验室将1944年发明的纸层析方法用于分开核苷酸、用紫外线分光光度仪检测核酸,提高了灵敏性与可靠性(Vischer and Chargaff, 1947, 1948)。几年内,他们很快发现A:G:C:T不等于1:1:1:1,推翻了单调重复的四核苷酸假说、支持Avery-MacLeod-McCarty提出的核酸可能具有化学特异性(Chargaff et al., 1949;Vischer, Zamenhof and Chargaff, 1949)。有了DNA酶以后,他们将牛胰DNA降解为多个片段,发现不同片段含A、G、C、T量不同,从而提出DNA链中核苷酸有非单调的排列(Zamenhof and Chargaff, 1950;Tamm et al., 1953)。
Chargaff最初只关注不同来源DNA中核苷酸是否不同,其实他们1949年的文章中的资料就还有更多信息,如A:T=1:1、G:C=1:1(Chargaff et al., 1949; Vischer, Zamenhof and Chargaff, 1949),但他们当时没有注意到。1950年,Chargaff写综述总结自己实验室的工作时,意识到多种来源的DNA中常常A:T=1:1、G:C=1:1(Chargaff,1950),但他不清楚是巧合还是真的规律,只提请注意。即使1951年他们在精子再度观察到这样的比例,也不敢断定是否巧合(Chargaffet al., 1951)。这一比例后人称为Chargaff规则,为Watson和Crick的DNA双螺旋两条链的碱基配对提供了伏笔。Chargaff还发现同种生物的DNA相同,而不同生物之间不同,说明DNA有生物特异性,有时这也被称为Chargaff规则之一。他还注意到同一生物的RNA在不同器官不同,他提出可能这些不同RNA参与器官分化(Chargaff,1950)。
9-4 转化的普遍性及其与遗传的关系
认同Avery-MacLeod-McCarty(1944)工作有很大意义的科学家也认为当时还不能完全接受DNA是遗传物质。因为即使转移因子只含DNA而无蛋白质,Avery-MacLeod-McCarty(1944)的结果还可以有多种解释,1958年的诺贝尔奖获得者在1956年总结为1)转化因子不是遗传物质,而是一种突变剂;2)多糖合成的自身催化剂;3)能够诱导宿主细胞荚膜合成反应的细菌病毒;4)细胞质基因或形态发生源;5)未进入细胞起作用;6)细菌遗传机构中能检测的一部分;7)特有的机制、无普遍性(Lederberg, 1956):。
1946年,McCarty已经知道转化因子的作用是可遗传的,被转化的细菌不仅性状传代,而且从后代提取到的转化活性可以高于最初用的转化活性,说明转化因子不仅能诱导荚膜多糖合成,而且能在细菌中自我复制,这些特性既与基因相似、也与病毒相似(McCarty, 1946b),所以Lederberg在1956年都没意识到他提的7点已有很多可以为1946年McCarty已知的结果所排除,而Lederberg还属于特别推崇Avery-MacLeod-McCarty者之一。
对于普遍性的问题,1947年法国的André Boivin报道可用来自一种抗原性的大肠杆菌的DNA转化另一大肠杆菌的抗原性,他称为定向突变(directed mutation)(Boivin, 1947);1949年Austrian和MacLeod在肺炎球菌可同时做到三个性状的转化(Austrian and MacLeod, 1949);1951年Hotchkiss发现从青霉素抵抗的细菌可得DNA,将青霉素抗药性转给其他细菌(Hotchkiss, 1951);1951年美国哥伦比亚大学儿科的Hattie Alexander与Grace Leidy发现流感嗜血杆菌的R类也可以被S类的DNA所转化,而且这时他们用了结晶纯的DNA酶,证明转移因子是DNA(Alexander and Leidy, 1951)。他们1953年发现流感嗜血杆菌的链霉素抗药性可通过DNA转化到原来对链霉素敏感的流感杆菌,起转化作用的DNA可被结晶纯的DNA酶所降解,他们还发现脑膜炎球菌分型相关的性状可通过DNA转化(Alexander and Redman, 1953)。
DNA可在多种细菌之间转化多种可以检测的性状,转化后的性状都可在代间遗传,从被转化的细菌的后代获得的DNA本身也有转化活性,证明DNA的作用之普遍性和可遗传性。
9-5 Hershey-Chase实验
应该说1946年McCarty和Avery文章后,DNA是转化因子的可能性非常大,而Hotchkiss(1952)纯化的转化因子蛋白质含量低于0.02%,1951年Alexander与Leidy用结晶纯的DNA酶可降解流感杆菌的转化活性就基本证明了DNA的转化因子。但如果要一直质疑,还可以说降解了DNA链后导致上面附着的微量蛋白质活性不稳定。要完全证明只有DNA是转化因子,需要确定某些基因的核苷酸序列、然后化学合成同样的序列、再用来转基因,而当时在知识上和技术上都不可能做到,只能依靠不断增强的外围证据。
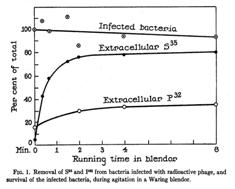
1950年代影响很大的工作是Hershey-Chase实验(Hershey and Chase, 1952)。Alfred Hershey(1908-1997)研究细菌的病毒(噬菌体),1950年到冷泉港实验室工作,Martha Chase(1927-2003)为其助手。噬菌体可以感染细菌,并在细胞内复制、细菌间传代。噬菌体外壳为蛋白质、内含DNA。Hershey和Chase用放射性同位素标记核酸和蛋白质以观察当细胞外的噬菌体感染细菌时,核酸和蛋白质的行为。磷32用于追踪DNA(所有DNA都含磷)、硫35用于追踪蛋白质(有些氨基酸含硫)。在噬菌体感染细菌几分钟后,Hershey和Chase摇动培养将细菌与细胞外的噬菌体分开,再检测磷和硫分别多少进入细胞、多少留在胞外,结果发现30%的磷32留在细胞外、80%的硫35留在细胞外;代间观察发现,30%的磷32传代、少于1%的硫35传代。这一实验被认为提供了很强的证据表明DNA是遗传物质、蛋白质不是。如果要比较,当然这些结果在纯度上远不如1946年MaCarty和Avery已经达到的程度,更不如Hotchkiss(1952)的程度。而且因为有些蛋白没有含硫的氨基酸,并非所有的蛋白质都能被硫35所标记。可能因为Hershey获诺贝尔奖的缘故,来自这一实验的图曾长年被很多教科书拷贝而未意识到其结论并非很好的证据。

9-6 细菌遗传学
Avery-MacLeod-McCarty后续研究的一个方面是以上验证DNA是转化因子、并推广其意义,另一方面是刺激以后的发展。
美国遗传学家Joshua Lederberg(1925-2008)于1945年念研究生的期间读到Avery-MacLeod-McCarty,他记录1945年1月20日一个晚上读文章后兴奋状态,认为意义“无限”(unlimited in its implications)、有突变特征、可以复制。他在1994年回顾1945年时认为Avery-MacLeod-McCarty可以声称:a)肺炎球菌有可遗传的性状,如血清学特异的多糖细胞荚膜,与致病性有关,并可在老鼠或血清中被选择;b)这些性状的遗传原基可通过无细胞的提取物在菌株之间转移,称为转化;c)转化因子的化学结构是DNA,不是蛋白质或其他大分子。如果这三点成立,那么就带来激进的概念:d)细菌有与高等动物一样的基因;e)基因是DNA;f)细菌可用于遗传学研究。
1943年,意大利裔美国犹太生物学家Salvador Luria(1912-1991)和德裔美国物理学家Max Delbrück(1906-1981)借助数学模型分析细菌对噬菌体抵抗性的统计规律,证明细菌可以自发出现遗传突变(Luria and Delbrück, 1943)。美国Stanford大学的Edward Tatum(1909-1975)1941年与遗传学家George Beadle(1903-1989)通过遗传突变研究粗糙链胞霉(Neurospora)的生化反应(Beadle and Tatum, 1941)、最终提出一个基因一个酶的概念。Tatum于1944年和1945年研究了细菌(K12大肠杆菌)的突变。
还在哥伦比亚大学念医学院的Lederberg从本科阶段就参加科学研究,其指导老师曾跟随Tatum用粗糙链胞霉。1945年1月,Lederberg从一位研究生那里获Avery-MacLeod-McCarty(1944)全文,读后兴奋不已。他想用链胞霉研究转化,结果发现他们最初用的链胞霉突变种很容易自发回复,没做成他预计的实验,但发表了他第一篇论文。他再转向研究细菌是否有“性”。他的老师听说Tatum要来东海岸到耶鲁工作,建议他到Tatum实验室。Lederberg给Tatum写信提出他的研究目的:细菌的性重组(sexual recombination)。他们在耶鲁很快出了结果,发现细菌的性生活:一个细菌将信息给另一细菌(Lederberg and Tatum, 1946; Lederberg, 1947)。细菌遗传学在1950和1960年代为遗传学的核心,其所用的细菌、发现的质粒(Lederberg,1952)等也成为分子生物学和1970年代诞生的生物技术产业的重要工具。
9-7解析DNA三维结构
美国生物学家James D Watson(1928-)是印第安纳大学Luria的研究生,他于1950年毕业,学了噬菌体和遗传学,他非常接受Avery-MacLeod-McCarty(1944)的观点,认为DNA就是遗传物质。Watson到英国剑桥大学以物理见长的、麦克斯韦曾为第一任主任的卡文迪许实验室做博士后期间,不愿按导师John Kendrew(1917-1997)安排研究蛋白质或病毒的结构,而与Max Perutz(1914-2002)的研究生Francis Crick(1916-2004)热衷讨论DNA的结构(Watson, 1968)。Watson和Crick与伦敦国王学院用X线衍射DNA的Maurice Wilkins(1916-2004)和Rosalind Franklin(1920-1958)有很多讨论,最初他们提出的三螺旋模型被Franklin指出有明显错误,其后还得益于看到Franklin拍摄的一张衍射图片,提出DNA双螺旋模型。Franklin也独立地提出了DNA双螺旋模型。1953年4月25日同一期Nature刊登(Watson and Crick, 1953a; Wilkins et al., 1953; Franklin and Gosling, 1953)。只有Watson和Crick(1953)提出了碱基配对,也就解释了A/T、G/C比例为何为一,很快他们提出DNA复制的机理(Watson and Crick, 1953b)。双螺旋结构很漂亮,而生物学意义重要的是碱基配对。物理学在分子生物学的建立过程起了很大作用。
注:
1)Avery-MacLeod-McCarty(1944)和Watson and Crick(1953)是二十世纪最重要的两项生物学研究。如果认为首先需要确定DNA是遗传物质,其后分析DNA结构才重要,也可以说Avery-MacLeod-McCarty更重要。
2)1944年,Avery已67岁,属很少见的在年龄很大的时候做出重要工作的科学家。MacLeod只有35岁,McCarty只有33岁。MacLeod一直是“少年得志”类,15岁高中毕业,23岁毕业于McGill大学医学院,到纽约大学任微生物系主任时仅32岁。
3)据Hammarsten的学生Reichard(2002)在研读解密的文件后介绍,1932年至1946年几乎每年Avery都因发现细菌多糖的抗原性而被提名诺贝尔奖。1946年后提名开始提到他的转化工作,但当时诺贝尔医学奖委员会懂核酸的关键成员Hammarsten虽首先提议委员会考虑这一工作,但他每次都认为不行,前几年是考虑到提取物易污染、难排除蛋白质起转化作用的可能性,等到1952年Hershey和Chase文章、1953年Watson和Crick文章后,他和其他成员相信DNA是转化因子,Hammarsten的评估报告认为DNA是转化因子,但其作用机理不明、所以得奖还早。1953年诺奖给三羧酸循环的发现者Krebs和辅酶A的发现者Lipmann、1954年奖给发现脊髓灰质炎病毒的Enders、Weller和Robbins。而1955年Avery去世后,DNA方面的诺贝尔奖都比较快,包括1959年发错了一半。
4)Kossel获1910年诺贝尔化学奖,因为“通过其对蛋白质的研究,包括核物质,对细胞化学知识的贡献”,委员会看重他研究蛋白质,而他的诺贝尔演讲前面大半内容是核酸。Lederberg于1946和1947年发表重要结果后,不再念哥伦比亚大学的医学院而转到耶鲁做研究生(Lederberg,1987),他的研究生工作使他在33岁与Beadle和Tatum共享1958年诺贝尔奖。Arthur Kornberg(1918-2007)因于1956年发现DNA多聚酶获1959年诺贝尔奖。Perutz与Kendrew因研究血红蛋白和肌球蛋白的三维结构获1962年诺贝尔化学奖。Watson与Crick获1962年诺贝尔生理或医学奖。Delbrück、Luria和Hershey获1969年诺贝尔奖。
5)法裔科学家René Jules Dubos多才多艺,1948年因研究土壤细菌及其抗生素与导师Waksman一道获得Lasker奖(Waksman因为发现链霉素获1952年诺贝尔奖,但他另一研究生Albert Schatz(1920-2005)认为自己是发现链霉素的主力)。Dubos被Avery招到洛克菲勒后,除几年在哈佛任教外都在洛克菲勒。他写了很多包括科普的书,其中1968年出版的《如此人性的动物》(So Human an Animal)一书获1969年普利策奖。Avery找到Dubos是巧遇,Dubos于1927年研究生毕业时,有人建议他到洛克菲勒求教法国同胞、1912年诺贝尔奖得主Alexis Carrel(1873-1944),他们到餐厅午餐时Dubos旁边正好坐着Avery,两人一拍即合,Avery说了他面临的问题(肺炎球菌荚膜多糖),Dubos说肯定可以从土壤细菌中找到降解的酶。
6)Kossel于1884年发现的组蛋白,在二十世纪末和二十一世纪初为分子生物学研究的热点。1974年,美国Stanford大学的Roger Kornberg提出DNA重复地、规则地环绕组蛋白形成核小体(Kornberg and Thomas,1974;Kornberg,1974)。美国洛杉矶加州大学(UCLA)的Michael Grunstein实验室通过酵母遗传学证明组蛋白和核小体对基因转录的重要性。1996年美国的David Allis实验室发现组蛋白乙酰转移酶后(Brownell et al.,1996),组蛋白修饰成为很多人研究的表观遗传学的核心问题之一。
7)发现肺炎球菌的 因为是犹太人,在1930年代已年迈后没有被希特勒得势后的虐犹者所放过,被剥夺教授职位和行医执照。当时看来只是犹太人受迫害,一大批科学家被整和出逃导致有辉煌科学历史的德国迅速落后。
因为是犹太人,在1930年代已年迈后没有被希特勒得势后的虐犹者所放过,被剥夺教授职位和行医执照。当时看来只是犹太人受迫害,一大批科学家被整和出逃导致有辉煌科学历史的德国迅速落后。
8)Dochez一战时入伍为少校,1919年到霍普金斯大学任教,1921年回纽约在哥伦比亚大学任教,长期是Avery的室友和朋友。其研究自1919年改为链球菌,1928年改为感冒病毒。
参考文献
Alexander HE and Leidy G (1951) Determination of inherited traits of H. influenzae by desoxyribonucleic acid fractions isolated from type-specific cells. Journal of Experimental Medicine 93:345-359.
Alexander HE and Leidy G (1953) Induction of streptomycin resistance in sensitive Hemophilus influenzae by extracts containing desoxyribonucleic acid from resistant Hemophilus influenzae. Journal of Experimental Medicine 97:17-31.
Alexander HE and Redman W (1953) Transformation of typespecificity of meningococci: Change in heritable type induce by type-specific extracts containing desoxyribonucleic acid.Journal of Experimental Medicine 97:797-806
Alloway JL (1932) The transformation in vitro of R pneumococci into S forms of different specific types by the use of filtered pneumococcus extracts. Journal of Exerimental Medicine 55:91-99.
Alloway JL (1933) Further observations on the use of pneumococcus extracts in effecting transformation of type in vitro. Journal of Experimental Medicine 57:265-278.
Altmann R (1889).  .Archiv fürAnatomie und Physiologie. Physiologische Abteilung. 524-536, Leipzig.
.Archiv fürAnatomie und Physiologie. Physiologische Abteilung. 524-536, Leipzig.
Amoss HL (1925) The composite nature of a pure culture of avirulent pneumococcus. Journal of Experimental Medicine 41:649-662.
Artwright JA (1921) Variation I nbacteria in relation to agglutination both by salts and by specific serum. Journal of Pathology and Bacteriology 24:36-60.
Astbury WT (1947) X-ray studies of nucleic acids. Symposium of the Society of Experimental Biology 1:66-76.
Astbury WT and Bell FO (1938) X-ray studies of thymonucleic acid. Nature 141:747-748.
Austrian R and MacLeod CM (1949) Acquition of M protein by pneumococci through transformation reactions. Journal of Experimental Medicine 89:451-460.
Austrian R and Gold J (1964) Pneumococcal bacteremia with especial reference to bacteremic pneumococcal pneumonia. Annals of Internal Medicine 60:759-776.
Austrian R, Douglas RM, Schiffman G (1976) Prevention of pneumococcal pneumonia by vaccination. Transactionsof the Association of American Physicians 89:184-189.
Avery OT, Chickering HT, Cole R,Dochez AR (1917) Acutelobar pneumonia: prevention and serum treatment. Monograph No. 7, New York: Rockefeller Institurte for Medical Research.
Avery OT and Dubos R (1931) Protective action of a specific enzyme against type III pneumococcus infection in mice. Journal of Experimental Medicine 54:73-90.
Avery OT and Goebel WF (1933) Chemoimmunological studies on the soluble specific substance of Pneumococcus: I. the isolation and properties of the acetyl polysaccharide of Pneumococcus type I. Journal of Experimental Medicine 58:731-755.
Avery OT, MacLeod CM and McCarty M (1944) Studies on thechemical nature of the substance inducing transformation of pneumococcal types.Journal of Experimental Medicine 79: 137-158.
Bernal JD and Crowfoot DM (1934) X-ray photographs of crystalline pepsin. Nature 133:794-795.
Beadle GW, Tatum EL (1941) Genetic control of biochemicalreactions in Neurospora. Proceedings of the National Academy of Sciences 27:499-506.
Boivin A (1947) Directed mutation in colon bacilli, by aninducing principle of desoxyribonucleic nature: its meaning for the general biochemistry of heredity. Cold SpringHarbor Symposium on Quantitative Biology 12:7-17.
Brachet J (1942) La localization des acides pentosennucléiquesdans les tissus animaux et les oeufs d’Amphibiens en voie de développeent. Archiv de Biologie 53:207-257.
Brown R (1833) Obesrvation on organs and mode offecundation in Orchideae and Asclepiadeae. Transactions of the Linnean Societyof London 16:685-716.
Brownell JE, Zhou J, Ranalli T, Kobayashi R, Edmondson DG,Roth SY, Allis CD (1996). Tetrahymena histone acetyltransferase A: a homolog to yeast Gcn5p linking histoneacetylation to gene activation. Cell 84:843-851.

Caspersson T, Hammarsten E and Hammarsten H (1935).Interactions of proteins and nucleic acid. Transactionsof the Faraday Society 31:367-389.
Caspersson T and Schultz J (1938) Nucleic acid metabolism of the chromosomes in relation to gene reproduction. Nature 142:294-295.
Caspersson T and Schultz J (1939). Pentose nucleotides in the cytoplasm of growing tissues. Nature143:602.
Chargaff E (1950) Chemical specificity of nucleic acids and mechanism of their enzymic degradation. Experientia 6:201-209.
Chargaff E and Zamenhof S (1948) The isolation of highly polymerized dexoxypentose nucleic acid from yeast cells. Journal of Biological Chemistry 173:327-335.
Chargaff E, Vischer E, Doniger R, Green C and Misani F(1949) The composition of the desoxypentose nucleic acids of thymus and spleen.Journal of Biological Chemistry177:405-416.
Chargaff E, Lipshitz R, Green C and Hodes ME (1951) The composition of the deoxyribonucleic acid of Salmon sperm. Journal of Biological Chemistry 192:223-230.
Dahm R (2005). Friedrich Miescher and the discovery of DNA.Developmental Biology 278:274-288.
Dahm R (2008). Discovering DNA: Friedrich Miescher and the early years of nucleic acid research. Human Genetics 122:565-581.
Dale HH (1946) Address of the president. Proceedings of the Royal Society B 133:123-139.
Darnell J (2011) RNA, life’s indispensable molecule. pp.43-44, Cold Spring Harbor Laboratory Press, New York.
Dawson MH (1928) The interconvertibility of “R” and “S”forms of pneumococcus. Journal of Experimental Medicine 47:577-591.
Dawson MH (1930) The transformation of pneumococcal types:II the interconvertibility of type-specific S pneumococci. Journal of Experimental Medicine 51:123-147.
Dawson MH and Sia RHP (1931) In vitro transformation of pneumococcal types. I. A technique for inducing transformation of pneumococcal types in vitro. Journal of Experimental Medicine 54:681-699.
Dobzhansky T (1941) Genetics and the origin of the species, p. 47, Columbia University Press, New York.
Dochez AR and Avery OT (1917) The elaboration of specific soluble substance by pneumococci during growth. Journal of Experimental Medicine 26:477-493.
Dochez AR and Gillespie LJ (1913) A biologic classificationof pneumococcus by means of immunity reactions. Journal of the American Medical Association 61:727-732.
Dochez AR and Avery OT (1916) Antiblastic immunity. Journal of Experimental Medicine23:61-68.
Downie AW (1972) Pneucococcal transformation-a backward view: Fourth Griffith Memorial Lecture. Journalof General Microbiology 73:1-11.
Feulgen R and Rossenbeck H (1924) Mikroskopisch-chemischerNachweis einer Nucleinsure von Typus der Thyomonucleinsure and die darauf beruhendeelective Frbung von Zellkernen in microskopicschen Praparaten. Hoppe-Seyler’s Z. Physiol. Chem. 135: 203-248.
Franklin R and Gosling RG (1953) Molecular configuration insodium thymonucleate. Nature 171:740-741.
Goebel WF, Avery OT, Babers FH (1934). Chemoimmunological studies on conjugated carbohydrate proteins. IX. The specificity of antigens prepared by combining the p-aminophenol glysides of disaccharides with protein.Journal of Experimental Medicine60:599-617.
Griffith F (1922) Types of pneumococci obtained from cases of lobar pneumonia. In Reports on Publicand Medical Subjects, № 13, pp. 20-45, His Majesty’s Stationery Office,London.
Griffith F (1923) The influence of immune serum on the biological properties of pneumococci. In Reportson Public and Medical Subjects, № 18, pp. 1-13. His Majesty’s StationeryOffice, London.
Griffith F (1928) The significance of pneumococcal types. Journal of Hygiene 27:113-159.
Heidelberger M and Avery OT (1923) The soluble specific substance of Pneumococcus. Journal of Experimental Medicine 38:73-79.
Henriques-Normark B and Tuomanen EI (2013) The pneumococcus: epidemiology, microbiology and pathogenesis. Cold Spring Harbor Perspective doi:10.1101.
Hershey AD and Chase M (1953) Independent functions ofviral protein and nucleic in the growth of bacteriophage. Journal of General Physiology 36:39-56.
Hotchkiss RD (1951) Transfer of penicillin resistance inpneumococci by the desoxyribonucleate derived from resistant cultures. Cold Spring Harbor Symposium on QuantitativeBiology 16:457-461.
Hotchkiss RD (1952) The biological nature of the bacterial transforming factors. Experimental Cell Research Supplement 2:383-389.
Hotchkiss RD (1965) Oswald T Avery: 1877-1955. Genetics 51:1-10.
Itakura K, Hirose T, Crea R, Riggs AD, Heyneker HL, BolivarF, Boyer HW (1977) Expression in Escherichia coli of a chemically synthesized gene for the hormone somatostatin. Science 198:1056-1963.
Jones ME (1953) Albrecht Kossell, a biographical sketch. Yale Journal of Biology and Medicine 26:80-97.
Judson H (1979) The eighth day of creation: makers of therevolution in biology. Simon and Schuster.
Kasten FH (2003) Robert Feulgen and his histochemicalreaction for DNA. Biotechnic and Histochemistry 78:45-49.
Kornberg R and Thomas JO (1974). Chromatin structure: oligomers of the histones. Science 184:865-868.
Kornberg R (1974). Chromatin structure: a repeating unit ofhistones and DNA. Science184:868-871.
Kossel A (1879). Ueber das Nuclein in der Hefe I. Zschar Physiol Chem 3:284-291.
Kossel A (1891)Ueber die chemische Zusammensetzungder Zelle. DuBois-Reymond’s Arch181:181-186.
Kunitz M (1940) Crystalline ribonuclease. Journal of General Physiology 24:15-32.
Kunitz M (1948) Isolation of crystalline desoxyribonuclease from beef pancreas. Science 108:19-20.
Kunitz M (1950) Crystalline desoxyribonuclease. Journal of General Physiology 35:349-377.
Lederberg J (1947) Gene recombination and linked segregations in Escherichia coli. Genetics 32:505-525.
Lederberg J (1952) Cell genetics and hereditary symbiosis. Physiological Reviews 32:403-430.
Lederberg J (1956) Genetic transduction. American Scientist 44: 264-280.
Lederberg J (1994) The transformation of genetics by DNA:an anniversary celebration of Avery, MacLeod and McCarty (1944). Genetics 136:423-426.
Lederberg J (1987) Genetic recombination in bacteria: a discovery account. Annual Review of Genetics 21:23-46.
Lederberg J and Tatum EL (1946) Gene recombination in Escherichia coli. Nature 158:558.
Levene PA (1909) Ueber die Hefenucleinsaeure. Bhichem Zeitschr 17:120-121.
Levene PA and Jacobs WA (1912). On the structure of thymusnucleic acid. Journal of Biological Chemistry 12: 411-420.
Levene PA (1917) The structure of yeast nucleic acid. Journalof Biological Chemistry 31: 591-598.
Levene PA (1919) The structure of yeast nucleic acid: IV.ammonia hydrolysis. Journal of Biological Chemistry 40: 415-424.
Levene PA (1920a) The structure of yeast nucleic acid: V. ammoniahydrolysis. Journal of Biological Chemistry 41:19-23.
Levene PA (1920b) The structure of yeast nucleic acid: ammonia hydrolysis: the so called trinucleotide of Thannhauser and Dorfmüller. Journal of Biological Chemistry 43:379-382.
Levene PA and London ES (1929) The structure of thymonucleic acid. Journal of Biological Chemistry 83:793-802.
Liu S and Wu H (1938) Mechanism of the recovery of antibody recovery from immune precipitate. Chinese Journal of Physiology 13:449-462.
Luria SE, Delbrück M (1943)Mutations of bacteria from virus sensitivity to virus resistance. Genetics 28:491–511.
McCarty M (1945) Reversible inactivation of the substance inducing transformation of pneumococcal types. Journal of Experimental Medicine 81:501-514.
McCarty M (1946a) Purification and properties of desoxyribonuclease isolated from beef pancreas. Journal of General Physiology 29:123-139.
McCarty M (1946b) Chemical nature and biological specificity of the substance inducing transformation of pneumococcal types. Bacteriology Reviews 10:63-71.
McCarty M and Avery OT (1946a) Studies on the chemicalnature of the substance inducing transformation of pneumococcal types. II. Effect of desoxyribonucleose on the biological activity of the transforming substance. Journal of Experimental Medicine 83:89-96.
McCarty M and Avery OT (1946b) Studies on the chemical nature of the substance inducing transformation of pneumococcal types. III. An improved method for the isolation of the transforming substance and its application to pneumococcus types. Journal of Experimental Medicine 83:97-104.
MacLeod CM, Hodges RG, Heidelberger M, Bernhard WG (1945)Prevention of pneumococcal pneumonia by immunization with specific capsular polysaccharides. Journal of Experimental Medicine 82:445-465.
Miescher JF (1871) Ueber die chemische Zusammensetzung der Eiterzellen. Medisch-chemische Untersuchungen 4: 441-460.
Miescher JF (1874) Die Spermatozoeneiniger Wirbelthiere. Ein Beitrag zur Histochemie. Verhandlungen der Naturforschenden Gesellschaft in Basel 6: 138-208.
Mirsky AE (1951) Some chemical aspects of the cell nucleus, in Leslie C. Dunn, ed., Genetics in the 20th Century, pp.127-153, New York.
Mirsky AE, Pollister AW (1942)Nucleoproteins of cell nuclei. Proceedingsof the National Academy of Sciences 28:344-352.
Mirsky AE, Pollister AW (1946)Chromosin, a desoxyribose nucleoprotein complex of the cell nucleus. Journal of General Physiology 30:117-148.
Mirsky AE, Ris H (1947) The chemical composition of isolated chromosomes. Journalof General Physiology 31:7-18.
Mirsky AE, Ris H (1949) Variable and constant components of chromosomes. Nature163:666-667.
Mirsky AE and Ris H (1951) The desoxyribonucleic acid content of animal cells and its evolutionarysignificance. Journal of General Physiology 34:451-462.
Morgan TH (1910) Sex-linked inheritance in Drosophila. Science32:120-122.
Morgan AH, Sturtevant AH, Muller HM, and Bridges CB (1915) The mechanisms of Mendelian heredity. Henry Holt & Co., New York.

Reichard P (2002) Oswald T Avery and the Nobel Prize in medicine. Journal of Biological Chemistry 277:13355-13362.
Reimann HA (1925) Variations in specificity and virulence ofPneumococci during growth in vitro. Journal of Experimental Medicine 41:587-600.
Reimann HA (1929) The reversion of R to S Pneumococci. Journal of Experimental Medicine 49:237-249.
Schultz J and Caspersson T (1940)Ribonucleic acids in both nucleus and cytoplasm and the function of the nucleolus. Proc Natl Acad Sci USA26:507-515.
Schultz J (1941) The evidence of the nucleoprotein nature of the gene. ColdSpring Harbor Symposium on Quantitative Biology 9:151-167.
Schultz J (1943) Physiological aspectsof genetics. Annual Review of Physiology 5:35-62.
Sevag MG (1934) Eine neuephysikalische enteiweissungemethode zur darstellung biologisch wirksamer subsanzen. Biochem Z 273:419.
Sia RHP (1926) Presence of type specific pneumocco-opsonins in sera of animals naturally resistant to pneumococcus infection. Experimental Biology and Medicine 24:709-711.
Sia RHP and Dawson MH (1931) In vitro transformation of pneumococcaltypes II. The nature of the factor responsible for the transformation ofpneumococcal types. Journal ofExperimental Medicine 54:701-710.
Signer R, Caspersson T, Hammarsten E(1938) Molecular shape and size of the thymonucleic acid. Nature 141:122.
Stryker LM (1916) Variations in the pneumococcus induced by growth in immune system. Journal of Experimental Medicine 24:49-68.
Tamm C, Shapiro HE, Lipshitz R and Chargaff E (1953) Distribution density of nucleotides within a desoxyribonucleic acid chain. Journal ofBiological Chemistry 203:673-688.
Thomas R (1992) Molecular genetics under an embryologist’s microscope: Jean Brachet, 1909-1988. Genetics 131:515-518.
Thompson RHS and Dubos RJ (1938) The isolation of nucleic acid and nucleoprotein fractions from pneumococci. Journal of Biological Chemistry 125:65-74.
Vischer E and Chargaff E (1947) The separation andcharacterization of purines in minute amounts of nucleic acid hydrolysates. Journal of Biological Chemistry 168:781-782.
Vischer E and Chargaff E (1948) The separation andquantitative estimation of purines and pyridines in minute amounts. Journal of Biological Chemistry 176:703-714.
Vischer E, Zamenhof S and Chargaff E (1949) Microbialnucleic acids: the desoxypentose nucleic acids of avian tubercle bacilli and yeast. Journal of Biological Chemistry177:429-438.
Watson JD (1968) Double helix. Atheneum, New York.
Watson JD and Crick FHC (1953a) A structure for deoxyribosenucleic acid. Nature 171:737-738.
Watson JD and Crick FHC (1953b) Genetical implications of the structure of deoxyribonucleic acid. Nature 171:964-967.
Wilkins MHF, Stokes AR and Wilson HR (1953) Molecular structure of deoxypentose nucleic acids. Nature 171:738-740.
Wright AE, Morgan WP, Colebrook L, Dodge RW (1914).Observations on prophylactic inoculation against pneumococcus infections and on the result which have been achieved by it. Lancet 1:1-10, 87-95.
Zamenhof S and Chargaff E (1950) Dissymmetry in nucleotide sequence of desoxypentose nucleic acids. Journalof Biological Chemistry 187:1-14.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}