生物界罕见如孟德尔般做出超出时代、超出其他人很多的工作;更常见是在很多人工作的基础上做出渐变性的工作、偶尔带来“跃迁”。所以,非天才、有时甚至相当平凡的人在生物学可以做出很有意义的工作。
自然界极为罕见连体人(图1);十九世纪末实验胚胎学的诞生,却使人类在实验室得到连体动物;二十世纪末分子生物学的推进,一个学生一天可以注射特定分子制造成千的连体蝌蚪(图1)。

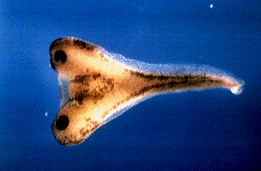
图1 连体现象
左,连体人(华裔美籍泰人Chang and Eng Bunkers, 1811-1874);右,连体蝌蚪(来自宾州大学Peter Klein)
连体胚胎反映了什么的生物学问题?研究它有什么意义?
发育生物学是研究一个细胞如何变成一个成体,并进一步变化直到死亡,包括细胞分裂、分化、生长、存活,组织和器官的形成和再生,个体的发育和衰老。发育生物学以前主要是研究胚胎发育,现在也研究超出胚胎基础过程的干细胞。而常为人类关心的组织、器官再生问题,缺乏基础研究的突破、有应用价值的结果更是微乎其微。发育生物学研究方法以前为简单的切割等操作,现代更多用分子生物学、也与物理和化学交叉。
1 预成论与后成论
动物的胚胎如何发育,曾有预成论(preformation)与后成论(epigenesis)的长期争论。预成论认为卵细胞(或精子、或受精卵)从开始就已形成了动物的各个部分,发育只是展开已有结构;而后成论认为受精卵并无成年的结构,而是在发育过程中逐渐形成。
希波克拉底(Hippocrates of Cos,公元前460-370)曾有类似预成论的观点:“胚胎中所有的都是同时形成。所有的肢体同时相互分开并生长,无一在其他之前或之后,而是那些自然较大的出现于较小的之前,并非更早形成”(引自Needham,1959)。
亚里士多德(Aristotle,公元前384-322)将预成与后成作为解释胚胎发育的两种对立概念。在《动物形成》一书中,他写道:“要么所有身体部件,如心、肺、肝、眼及其它,皆同时形成、或逐渐形成”。亚里士多德反对预成论:“前者不对是显而易见的,因为有些部件在胚胎已经清晰可见,而有些并非如此,且不是因为它们太小,比如肺比心大,但其发育出现晚于心……,那么就很明显,虽然有些东西制造部件,但非确定的物体,也不在精子中完整存在”。他支持后成论:“原则上,自然界一件东西来源于另外一件潜在可以成为终产物的东西……”。
后成论的支持者包括英国的哈维(William Harvey,1578-1657)、法国的笛卡尔(Rene Descartes,1596-1650)等,预成论的支持者包括意大利的Marcello Malpighi(1628-1694)、荷兰的列文胡克(Antonie van Leeuwenhoek,1632-1723)、德国的莱布尼茨(Gottfried Wilhelm von Leibniz,1646-1716)等。哈维在1651年称:卵中并无未来胚胎的任何部件,但潜在含有所有部件…有些部件先形成、有些后形成,都继续生长…这样我们称为后成(Needham,1959)。Malpighi于1673年再启争论,到18世纪预成论被认为定论(Meyer,1931;Needham,1959)。荷兰显微镜专家NicolaasHartsoeker(1656-1725)设想如果在显微镜下可以看到精子显示人体所有部分,可能的画面如图2。此图在被误认为是他的观察、而非其设想的情况下广为传播(Hill, 1985)。如果精子就有整个人体,那么发育不过是长大,而无需形成新的结构,甚至早先的人(比如传说的亚当或夏娃)应该含有所有现代人类的个体。当时预成论广为接受,争论是预成于卵子、还是精子。

图2 想象精子中的小人
Nicolaas Hartsoeker (1656-1725)曾于1694年描绘如果精子中有预成的小人可能的样子,此图常被误传为实际观察。
德国的CasparFriedrich Wolff(1733-1794)观察鸡胚发育。他于1759年在“发生理论”的论文中重提后成论,并叙述鸡胚在原本匀质的表面逐渐形成血管,1768年在“小肠形成”一书中描述鸡胚的肠子从一片组织折叠形成,所以血管和小肠皆非预先形成,而是逐渐形成。Wolff的结果推翻了传统的预成论(Meyer,1931;Needham,1959)。
2 自分化与依赖性分化
十九世纪后期,德国科学家Wilhelm Roux(1850-1924)区分两种观点:“不同部分的自分化”、“不同部分的分化性相互作用”。
Roux称自己的研究为“发育成因学”(Entwicklungsmechanik)(Sander,1991a)。以前的科学家包括他的老师Ernst Haeckel(1834-1919),对胚胎的研究依赖于观察,在描述的基础上提出理论;而德国的Oskar Hertwig(1849-1922)、法国的Laurent Chabry(1855-1893)等开始了实验研究(Fischer, 1990; Sander,1991a);Roux提倡分析性实验,不仅要有实验结果、还要思考结果、检验理论(Sander,1991a;Hamburger, 1997)。他们开创的学科到二十世纪被称为“实验胚胎学”(Experimental Embryology)。
Roux选择了重要“自分化”还是“依赖性”分化作为研究的问题,实验模型用两栖类胚胎。1888年,他发表了对蛙胚研究的结果和结论。他集中研究受精卵第一次分裂后成为的两个细胞是自主发育、还是相互依赖。为此,他在受精卵分裂后,用加热的针“杀死”其中一个细胞,观察未受针刺的细胞如何发育,他观察到剩下的细胞本身继续分裂、发育后形成半个胚胎(图3),据此Roux认为最初两个细胞之一的失去并不影响另外一个细胞的发育,所以两个细胞发育是独立的(Roux,1888)。


图3 Roux的热针杀死细胞实验
上排为示意图,下排为Roux的结果,引自Sander,1991b。
德国的HansDriesch (1867-1941)曾于1887年和1889年师从Haeckel。为了验证Roux的结论,Driesch在意大利Naples海洋生物实验站用海胆胚胎做实验。此前已知摇晃海胆的受精卵可以出现脱落的情况,Driesch在海胆受精卵分裂到两个细胞的时期,摇晃海胆几分钟,有些海胆胚胎的两个细胞会分开,Driesch将它们分别培养,然后观察分开的两个细胞如何发育。与Roux结论的预计不同,分开的两个细胞分别形成两个完整的、虽然体积只有正常一半大小的胚胎。如果在卵受精后分裂两次变成四个细胞时再摇晃,四个细胞都分别独立形成完整的胚胎(Driesch,1891)(图4)。这些结果显示了“调节”现象:胚胎的部分可以变成整体。对调节的解释说明:在胚胎两细胞期(或四细胞期),每个细胞都具有形成完整胚胎的潜力,但它们的相互作用导致每个细胞在其他细胞存在的情况下分别只参与形成半个胚胎(或四分之一个胚胎)。也就是说,细胞之间一定有相互作用。
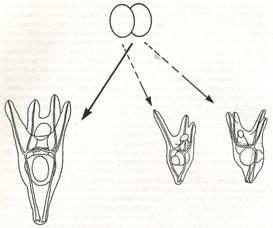
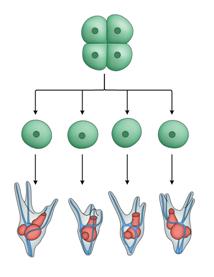
图4 Driesch的海胆胚胎细胞分开培养实验
左为两细胞期分开实验的示意图(自Hamburger,1988);右为四细胞期分开的示意图。
Roux和Driesch等创造了发育生物学的第一个高峰。Roux启迪了思路、开创了途径,Driesch有更严格的实验和分析。Roux未接受Driesch结果的重要性,认为低等的无脊椎动物海胆所得到的结论不如脊椎动物可靠。
德国的Hertwig(1893)最早试图用丝线分开两细胞期蛙胚的两个细胞(constriction,结扎实验),未成功(Beetschen, 1996)。美国的Morgan(1895)、意大利的Herlitzka(1895)、德国的Endres (1896)等曾有类似工作。
德国的HansSpemann(1869-1941)在Würzburg大学随Theodor Boveri做研究生,1895年获博士学位。Spemann在Hertwig、Endres等基础上,在蝾螈受精卵第一次分裂后,用头发把两个细胞分开,结果两个细胞可以分别独立地形成完整的蝌蚪,证明脊椎动物胚胎的细胞也有相互作用(Spemann,1901,1902,1903)(图5左)。
Spemann发现结扎是否导致两个完整胚胎形成取决于胚胎第一次细胞分裂的平面(图5中和右)。如果分裂平面与胚胎未来的左右相应,分开的两个皆完整胚胎。如果分裂平面与背腹相应时,背侧部分可以形成一个几乎完整的胚胎(含神经系统等重要结构)、而腹侧部分只能形成皮肤等结构而不形成完整的胚胎,说明胚胎具有腹侧和背侧的不对称性。Spemann认为腹侧缺乏某种“分化物质”:这种分化物质可能是器官形成或释放所需的未组织的物质、也可能是有组织的胚胎物质具有分化为各种器官的能力、或者引导其它细胞分化(Spemann,1901; Hamburger,1988)。
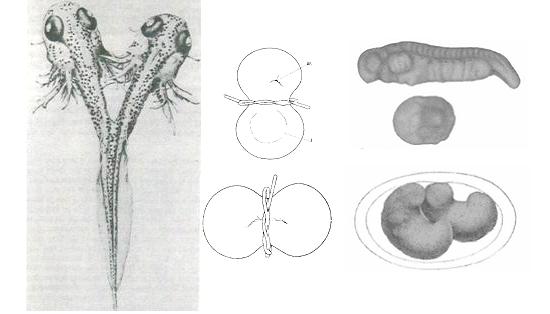 |
图5 Spemann的结扎实验
左为Spemann于1903年获得的结果(来源于Spemann,1903);中上示意当分裂的平面真好在身体左右之间,右上为中上所示切分方式的结果;中下示意分裂平面为前后而非左右,右下为中下所示切分方式的结果(来源于Spemann,1924)。
3 晶状体诱导
器官中不同部件优美地配合,既是进化的奇妙、也是发育的奥秘。不同部件是否在发育中有相互关系?
例如:美妙的眼睛中,晶状体(lens)在解剖上位于视网膜前特定部位、功能上调节光线准确地投射到视网膜。那么在发育过程中,晶状体与视网膜的发育前体结构视杯(optic cup,亦称optic vesicle)是否相关?比如空间、时间的协调,抑或有依赖关系?
1901至1912年,Spemann曾研究晶状体形成,认为在发育早期,视杯诱导晶状体产生。支持诱导机理的实验包括:Spemann于二十世纪初报道在去除视杯后,晶状体不能形成(Spemann, 1901b);美国霍普金斯大学的WarrenLewis(1870-1964)发现将原本不形成晶状体的外胚层移到视杯上方,可以形成晶状体(Lewis,1904);Lewis还发现将视杯移植到其他部位,导致新部位形成晶状体(Lewis,1906)(图6)。
其他研究者以及Spemann本人还看到有些胚胎晶状体产生不依赖视杯,当时以为有晶状体依赖视杯、和晶状体不依赖视杯的两类胚胎。1980年代后,视杯诱导晶状体的简单方式不被认可,而更复杂(Grainger et al.,1988)。
对于Spemann时代的胚胎学家来说,晶状体的研究为胚胎诱导的研究埋下了伏笔。
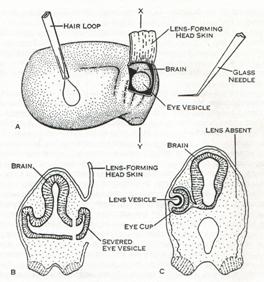
图6 晶状体诱导
上图示晶状体形成区域与眼杯区域的空间关系和晶状体诱导实验的工具;
下左示切除眼杯;
下右示眼杯切除后,晶状体也不能形成
(来自Hamburger,1988)。
4 移植实验
两栖类胚胎发育最初从一个受精卵、分裂到几千个细胞皆为裂殖胚(blastula,所含的每个细胞称为blastomeres),其后肠胚(gastrula)。在动物极与植物极交界的附近,细胞开始内卷(invagination),为肠胚形成的开始。内卷首先从背侧开始、继而侧面、后在腹侧,最后形成一圈,为裂殖孔(blastopore),背侧内卷时偏动物极的上方部位称为“背唇”(thedorsal blastopore lip,简称the dorsal lip)(图7)。
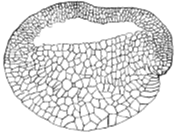
图7 背唇
左图为蛙发育的肠胚初期背唇照片,自Peter Klein;
右图为Morgan(1905)示意的截面图。
十九世纪Roux与Driesch等实验对象为裂殖胚。Spemann将结扎实验延伸到不同的胚胎发育期,发现一直到肠胚期如果分开背腹部分都有差别。分离的腹侧部只能形成皮肤、和腹侧及侧面中胚层组织入血液和间充质等,而分离的背侧部产生神经系统、和背侧的中胚层结构如脊索和体节。在分离时,背侧部分含背唇。
Roux和Morgan认为胚胎早期赤道部分对发育很重要,Roux还认为背唇可以产生背侧的外胚层结构神经管(Roux,1892; Sander, 1991b)。
1907年,霍普金斯大学的Lewis将肠胚期的背唇从一个供体胚胎移植到发育更晚期的宿主胚胎,结果发现宿主胚胎因为移植而多出现背侧中胚层组织脊索和肌肉、及外胚层组织神经管(图8)。Lewis认为:来自供体胚胎的背唇被移植到宿主的后自主形成了中胚层和外胚层组织,因为背唇的发育命运已经决定了(determined)(Lewis,1907),也就是认为背唇自主发育为脊索、肌肉和神经管,没有提出其中是否有因为背唇诱导宿主而产生的结构。
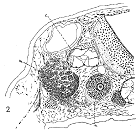
图8 背唇移植的后果
肠胚期背唇移植到tailbud期蛙胚7天后,观察到脊索(c)、肌肉(m)和神经(n)组织。来自Lewis,1907。
1916年,Spemann利用常见蝾螈(Triturustaeniatus)每个胚胎着色的自然差别,在两个同种蝾螈胚胎之间进行移植,通过着色深浅跟踪移植的供体在宿主胚胎中的发育命运。他发现肠胚期神经前体区域如果移植到另一胚胎的表皮区域,会形成表皮,而肠胚的表皮区域移植到神经区域会形成神经,都不按供体原有命运,而按被移植到的宿主区域改变了发育命运,说明在移植前它们的命运没有固定,而可以为新邻居所影响再确定(Spemann,1916,1918,1919)。
Spemann用蝾螈重复了Lewis蛙的背唇移植实验,获得同样结果。最初记录在他1916年4月20日的笔记中(Fäler,1996)。但他和Lewis一样,以为供体的背唇直接成为脊索和神经组织,而未意识到出现了诱导现象。Spemann在1918年的文章中还标明第二根神经管来源于移植的背唇(Spemann, 1918;Sander and Fässler,2001)(图9)。
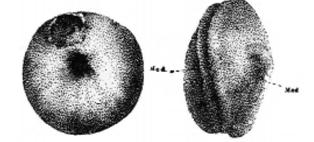
图9 背唇移植导致第二根神经管形成
Spemann(1918)认为背唇本身直接成为了脊索和神经管。来自Sander and Fässler(2001)
Spemann出现这样错误的原因是他不清楚背唇的命运,以为肠胚期的背唇两层分别形成脊索和神经板(Hamburger, 1988; Fäler,1996)(图10),他称背唇为“分化中心”(Spemann,1918)。
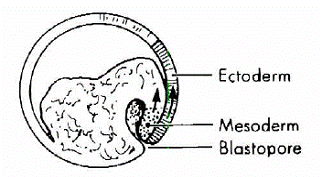
图10 背唇的发育命运
Spemann误认为背唇内层的命运为中胚层,背唇外层为外胚层,自Hamburger,1988。
如果Spemann仔细分析自己在1918年发表的结果(图11),可以看到:神经管来源的区域在背唇以上的区域。

图11 神经管的发育来源
Spemann(1918)结果显示移植蝾螈肠胚期背唇以上的区域形成神经管。
1919年海德堡大学的Hans Petersen在读到Spemann(1918)后,去信Spemann指出背唇的表层也应该是中胚层(脊索、体节)、而不形成神经管(Hamburger,1988;Sander and Fässler,2001),因此Spemann决定要研究背唇移植后导致的第二根背轴中脊索和神经管到底是来自移植供体、还是宿主。
Spemann于1916和1918年报道的移植是在同种胚胎之间,依赖供体和宿主同种的两个个体之间自然的色素差异,在分析时不太容易区分。为了确切区分背唇的命运,需要在颜色不同的不同种胚胎之间做背唇移植。1896年,德国犹太科学家GustavJacob Born(1852-1900)首先在异种胚胎间移植,他从一种蛙胚切下一块移植到另一胚胎上形成嵌合胚胎(chimera)(Born,1896)。1898年,美国耶鲁大学的Ross Harrison (1870-1959)用同样方法在不同种、不同色素的蛙之间进行过胚胎组织的移植(Harrison,1898),可以在移植后发育成的蝌蚪中区分来源于供体和宿主的组织。Spemann于1916年做同种移植(homeoplastic transplantation)、1917年开始做异种移植(heteroplastic transplantation)。Spemann用不同颜色的蝾螈之间进行移植,他常用色浅的Trituruscristatus和色深的Triturustaeniatus(或Triturusalpestris),以便区分供体和宿主来源的胚胎组织。
Hilde Mangold(1898-1924)上大学主修自然科学,她在Frankfurt大学听Spemann的学术报告后要求转学到Spemann所在的Freiburg大学,1920年进Spemann实验室做研究生(Fässlerand Sander,1996)。Spemann给她的第一个课题是研究水螅(hydra),但实验遇到问题、Spemann也不能解决,再给她的课题是Spemann自己做过、所以技术上应该没有问题:背唇移植。将来源于色浅的T cristatus的背唇移植到色深的T taeniatus(或T alpestris)。
在Mangold尚未开始移植实验的1921年1月,Spemann单独提交的文章写道:“也许整个移植的背唇属于…内中胚层…如果这样的话,从taeniatus胚胎来的背唇移植到cristatus宿主后,(中胚层的)脊索和体节应该属于前者,而(外胚层)的神经板属于后者”(Spemann, 1921;Sander and Fässler,2001)。
很可能是在初稿提交后Mangold的移植实验开始有了结果,Spemann将Mangold的一个结果于1921年5月作为后记补录在Spemann(1921)论文中(Spemann, 1921;Sander and Fässler,2001)。Mangold将色浅的T cristatus肠胚期背唇移植到色深的T taeniatus肠胚侧面,观察到第二个背轴产生。1921年,Spemann用了组织者(organizer)一词描述背唇:在它正常存在或者移植到的部位,对未分化材料创造一定方向和程度的“组织场”(Spemann,1921;Hamburger,1988)。
5 胚胎诱导与组织者
1921年和1922年,Mangold记录了259次移植实验,其中28个宿主胚胎出现第二背轴,当时培养胚胎的溶液欠佳、也无抗生素,所以胚胎在手术后的成活率不高。Hilde于1923年曾与丈夫Otto Mangold合作做移植实验(217个宿主,其中17个有第二背轴),Spemann没选Mangold夫妇合作获得的结果(Fässlerand Sander,1996)。
1924年,Spemann和Mangold发表合作的文章,Spemann选择详细分析了Mangold获得的六个胚胎移植的结果(Spemannand Mangold,1924)。
文章首先指出蝾螈肠胚化开始时,不同部位的决定性不同,通过移植可以检验,发现有些组织移植到其他部位后,其发育命运可以改变,这些工作Spemann于1918和1921已报道,而为Otto Mangold于1922年、1923年所验证和推广。然后,他们指出,Spemann(1918)就发现背唇很不一样,移植到形成上皮的地方,背唇继续按自己确定的命运发育,而移植后的区域出现第二个胚胎原基,含神经管、脊索和体节。Lewis(1907)有类似的发现,只是用肠胚后期的背唇移植到更老一些的胚胎其他部位,观察到它发育为神经组织和体节。
Spemann和Mangold提出发育命运确定在先的区域可以决定其他尚未确定命运区域的发育。他们用的例子是胚胎旋转的实验结果。蛙卵细胞有极性,着色深、较轻的为动物极,着色浅、较重的为植物极(图14),这一极性维持到胚胎发育过程中,精子从动物极进入卵细胞,动物半球主要贡献于外胚层和中胚层,植物半球主要贡献于内胚层。1918年Spemann的研究中曾将肠胚的动物极半球、植物半球分开,然后180度或90度旋转后再重合,结果认为植物极影响动物极的发育。由此,他们用了“组织中心”(organization center)的概念:胚胎中先决定的区域,再送出一定方向和一定量的决定性影响。
他们认为虽然Spemann(1918)发现了胚胎不同区域的决定性不同,但当时只在同种胚胎之间进行移植,不易区分移植供体与宿主,而异种移植更好,比如将T cristatus胚胎中命运确定的区域移植到T taeniatus(或T alpestris)胚胎中未定区域。
文章描述的六个移植为1921年的Um8b,和1922年Um25b、Um214、Um131b、Um83b、Um132b。文章只在讨论Um25时有文字介绍、而未显示Um214的图片,而多个图片显示了其他每个移植胚胎在多个部位的切片。
Um8b第二根神经管中很长部分来源于宿主、但有相当部分来源于供体,脊索来自供体;Um25b第二根神经管完全来自于宿主,而脊索完全来源于供体;Um214b的第二根神经管也完全来自于宿主,而脊索完全来源于供体;Um131b的第二根神经管多来自宿主、少数部位细胞来自供体,脊索和体节来自供体;Um83b的第二根神经管来自宿主,体节多数细胞来自宿主,脊索和部分体节来自供体;Um132b第二根神经管多数来自宿主(后2/3部分少数细胞来自供体,皆在神经管的底板,floor plate),脊索与部分体节来自供体(图12)。可能是因为Um25b的两张组织切片的图不特别好看,所以一般教科书不太用这一最能突出神经管可能由脊索诱导产生的例子,而常用Um132b为例,Um132的第二根神经管比较长,其胚胎在体外培养的时期也较长。
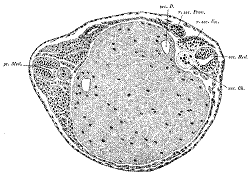
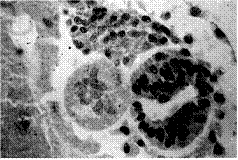
图12 Um132b第二根神经管的发育来源
左,Mangold原绘图,胚胎左边色深的第一根神经管和脊索、体节皆来源于宿主;右边第二根神经管的大部分细胞来自宿主,脊索来自供体,体节的部分细胞来自供体。自Spemann and Mangold, 1924。
右,后人拍摄Mangold留下Um132b原切片中第二根神经管及其附近结构的照片。第二根神经管在照片部分看来全是宿主细胞、无供体细胞,脊索全是供体细胞,体节有宿主和供体细胞。自Fässler & Sander, 1996。
在讨论中,Spemann和Mangold先谈了移植的方向、背唇的具体位置、发育期、第二原基内涵等。在机理讨论部分,他们明确提出肯定出现了诱导,但不清楚发生的确切的时间、是否直接诱导,提出两种可能:1)移植供体本身产生少量神经组织并存在于外胚层表面(如Um8b所见),而这些神经组织诱导宿主的细胞成为神经组织;2)中胚层诱导神经组织产生。他们当时的结果不能确定哪种解释为正确,提出了一些可能的实验。他们还定义和讨论了组织者的概念。
什么是组织者、组织者如何起作用可能在今天还有争论,而诱导为后来很多实验所证明,并不限于胚胎、也不限于发育,是细胞之间相互作用的一种范式。
6 余音缭绕
Walther Vogt(1888-1941)从1920年代初开始用染料跟踪胚胎细胞的命运(Vogt,1923),并于1925和1929年发表蛙胚的命运图(fate map)(Vogt, 1929)(图13)。他的图谱清楚地显示背唇本身只成为中胚层,Spemann以前以为背唇深层和浅层不同命运、浅层成为神经胚层的想法肯定不对。
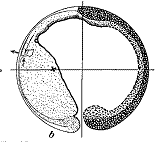
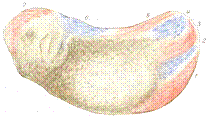
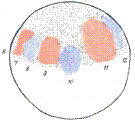
图13 Vogt的命运图谱
左,裂殖胚或肠胚早期多处放置活体染料;中,同左的胚胎到蝌蚪期,观察染料着色的区域成为何种组织、器官;右,总结背唇的命运为中胚层,而不贡献于神经板。自Vogt(1929)。
Spemann的研究生Johannes Holtfreter(1901-1992)、Otto Mangold等推进了神经诱导的研究,包括诱导的区域性、建立体外诱导模型等。Holtfreter实验室的中国留学生庄孝惠研究了宿主胚胎不同区域对外源性诱导物的反应(Hamburger et al.,1988)。
1919至1924年Spemann没做实验,1925年重回实验第一线,1929年开始用不同方法“破坏”背唇的结构(切、压、加热、冷冻、干燥),观察经过这些处理后,背唇是否还有诱导的活性(Hamburger, 1988),结果多为阴性。Holtfreter离开Spemann实验室后独立做过类似的研究,他改进了培养液,大大提高了胚胎的存活率,他发现多种方法处理后的背唇仍然有神经诱导的活性。1932年,Spemann与三个学生合作发表了一篇论文、四人分别写一段(Bautzmann et al.,1932),其中Holtfreter一人做过130个胚胎,其他人做过少量的胚胎:Hermann Bautzmann观察到背唇加温到60度处理5到10分钟后,还可能有神经诱导活性;Holtfreter发现背唇在60度干燥、100度加温、冷冻后,仍有神经诱导活性;Spemann发现背唇用96%的酒精处理3分钟后,仍有诱导活性,不过他只有一例胚胎;Mangold希望看诱导物质是否可弥散,将背唇附着于琼脂,干燥后将琼脂转到胚胎,检验其诱导性。Bautzmann等(1932)证明无需活着的背唇组织或细胞就可能有诱导活性,被认为开启了胚胎诱导分子的研究。
Spemann的研究生Else Wehmeier与化学家Fischer合作,试图用丙酮、乙醇、乙醚等提取背唇的神经诱导物质(Fischerand Wehmeier,1933)。剑桥大学的李约瑟(Joseph Needham,1900-1995)、Dorothy Moyle Needham(1896-1987)和Conrad Waddington(1905-1975)研究蝾螈提取物的诱导活性(Waddington et al.,1933; Needham et al., 1934)。但是,生化胚胎学(biochemical embryology)热潮中,并未得到诱导的分子。李约瑟转行研究中国科技,最终与一些华人和非华裔学者合作写出《中国科学与文明》。Waddington于1942年从原意为后成论的epigenesis衍生出epigenetics一词,当时他定义为“研究基因与其产物相互作用,导致表型产生的学科分支”,但基本涵义为发育,例如他于1952年出版《鸟的epigenetics》一书叙述鸡的胚胎发育过程、而非遗传学。Epigenetics经几重转用后成为今天的热门词汇“表观遗传学”。

图14 裂殖胚着色深的动物极,浅的植物极。自Peter Klein。
1940年代至1970年代的三十年为胚胎发育研究的寒冷期。其中最重要的是荷兰Utrecht大学的Pieter Nieuwkoop于1969年发现中胚层为内胚层诱导所产生。Nieuwkoop将裂殖胚从动物极到植物极分成几个部分,发现虽然动物极单独不能形成中胚层,但它如果与植物极共同培养,可以形成中胚层(Nieukwoop,1969a, 1969b,1970)。进一步通过同位素标记区分供体和宿主证明是植物极与动物极共培养时,中胚层来自动物极,所以是植物极诱导动物极产生中胚层(Nieukwoop and Ubbels,1972)。在正常发育过程中,中胚层之所以在动物极和植物极之间产生,是因为植物极发出信号诱导动物极和植物极之间的结构(Morgan时代称赤道区、后称边缘区marginalzone)形成中胚层。
由于背唇神经诱导曾被质问(Jacobson, 1982, 1983),英国两个实验室于1983年将辣根过氧化酶(horseradish peroxidase,HRP)注射到供体胚胎,在肠胚早期获得背唇,移植到同期宿主的腹侧,观察到第二根背轴,其中HRP只存在于脊索中,而神经管完全来源于宿主(Smith and Slack,1983;Gimlich and Cooke,1983),从而回答了质疑、再次证实了1924年的结论。
肠胚期胚胎的腹背轴已经形成,因此肠胚背唇移植导致背轴形成的实验需要在胚胎更早期进行研究。美国伯克利加州大学的John Gerhart实验室在64细胞期做移植实验,发现植物半球背侧的单个细胞如果移植到宿主胚胎的腹侧,可以诱导第二背轴产生(Gimlich and Gerhart,1984)。实际上,腹背轴的形成可以推早到受精:在受精前,卵细胞无腹背差别,而精子进入导致卵细胞内细胞骨架变化、引起一系列变化,致使精子进入一侧为将来胚胎的腹侧(Gerhart et al.,1989)。
所以,背唇移植实验延伸出中胚层诱导、腹背轴形成等多个重要问题的研究。
7 分子时代
1930年代没找到胚胎诱导相关的分子;1970年代重组DNA为代表的新技术导致分子生物学在生物医学多个领域的应用;1980至1990年代不同的方法帮助找到了中胚层诱导、背轴诱导和神经诱导的分子,并对神经诱导的机理有新的理解。
因为胚胎植物极有诱导中胚层的活性,哈佛大学的Douglas Melton通过mRNA差异性分布,找到非洲爪蟾卵细胞动物极和植物极分别特有(或富集)的mRNA(Rebagliati et al., 1985),其中植物极特有的Vg-1编码蛋白质为转化生长因子(TGF-)家族的成员(Weeks and Melton,1987)(图15)。美国旧金山加州大学(UCSF)的Kimelman和Kirschner用裂殖胚的动物极部分(所谓“动物帽”,animal cap)体外培养作为检测方法,筛查当时已知生长因子的诱导活性。动物帽单独培养只形成皮肤,而培养时如果存在成纤维细胞生长因子(fibroblast growth factor,FGF),可以出现中胚层(特别是腹侧和侧面的中胚层组织)(Kimelman and Kirschner,1987)。英国的Jim Smith发现爪蟾体外培养细胞系的分泌物有中胚层诱导活性(Smith, 1987),以后被证明为activin、也是TGF-家族成员。Activin的作用有剂量依赖性,低剂量诱导腹侧、侧面中胚层,高剂量诱导背侧中胚层(如背唇、或其衍化的脊索)。TGF-家族的骨形成蛋白(BMP,bonemorphogenetic protein)可以诱导腹侧中胚层产生。

图15 Vg-1的mRNA分布富集于植物极。自Rebagliati et al.,1994。
一个偶然的发现开启了对背轴形成分子机理的理解。研究癌基因的英裔美国分子生物学家Andrew McMahon与研究蛙胚的Randall Moon合作,将认为在老鼠过高或者异位表达导致乳腺癌的int-1基因所转录的mRNA注射到爪蟾受精卵,结果胚胎形成两个背轴 (McMahan and Moon,1989),这一意外的发现当时被认为是“分裂”了组织者,其后认为是激活了背轴形成的通路。从研究病毒导致老鼠癌症发现的int-1与从研究果蝇翅膀形成发现的wingless基因组成Wnt家族,产生分泌型蛋白,Wnt不同成员通过作用于其受体及信号转导分子参与多个生物学过程。它们在蛙胚参与背轴形成,但尚待确定胚胎内从哪一步开始用Wnt信号通路。1992年,伯克利加州大学的英裔美国科学家Richard Harland实验室发现表达于背唇的Noggin基因,编码分泌型蛋白质也能诱导背轴(Smithand Harland,1992)。
1993年,Harland实验室发现Noggin有神经诱导的活性(Lamb et al., 1993)。1994年,洛杉矶加州大学的阿根廷裔美国科学家DeRobertis实验室发现背唇表达Chordin、也编码有神经诱导活性的分泌型蛋白质(Sasaiet al., 1994)(图16)。

图16 背唇表达的Chordin 自Sasai et al.,1994。
Melton实验室的伊朗库尔德裔Ali Hemmati-Brivanlou在研究activin参与中胚层诱导的过程中,用activin受体制造的抑制分子阻止中胚层形成时,意外发现没有中胚层仍可出现神经系统,从而提出神经胚层的诱导在分子机理上是去抑制:胚胎内存在抑制神经胚层形成的分子,而如果抑制这些抑制性分子就可以“诱导”神经发生(Hemmati-Brivanlou and Melton, 1994)。抑制分子可能是BMP,而noggin和chordin等蛋白质可以直接结合BMP、并抑制BMP的活性(Piccolo et al., 1996;Zimmerman etal., 1996)。
8 总结
预成论与后成论的古老争论,在实验胚胎学兴起后,刺激了人们感兴趣研究早期细胞发育是完全独立自主、还是相互作用。
Spemann和Mangold(1924)的研究,实验设计类似于Lewis(1907)。Spemann曾于1916年、1918年重复Lewis的工作并沿用其理解,而Petersen于1919年向Spemann提出以前对背唇自分化命运的理解可能有问题,加上概念上曾有Spemann和Lewis等在二十世纪初研究晶状体可能被诱导产生的铺垫,导致Spemann决定采用Born(1896)和Harrison(1898)的方法,通过异种胚胎间移植确切地分清移植提供的背唇自身变成神经管、还是诱导宿主胚胎组织产生神经管,最终Spemann指导Mangold的实验获得结果提示神经管为背唇诱导所产生。
胚胎诱导的概念,不仅成为核心问题之一主导几十年的发育生物学研究,也影响了人们思考细胞相互作用所参与的其他生物学过程,但1930年代研究胚胎诱导的分子机理遭遇技术困难,直到1980、1990年代才获突破。
在实验胚胎学研究过程中多次发现不同方法可以产生连体胚胎,包括结扎实验、背唇移植实验、早期裂殖胚细胞移植实验、Wnt家族分子注射、以及本文未讨论的BMP受体抑制剂注射。这些研究帮助我们理解基本的发育生物学机理,但是,迄今并不清楚连体人的发生原因。


图17 Han Spemann与Hilde Mangold
左,1935年12月10日诺贝尔奖合影:物理得奖者居里夫人的女儿Irène Juliot-Curie、女婿Frédéric Joliot,生理学获奖者Spemann;
右,Hilde Mangold与幼子。
注1:预成论与后成论部分参考了作者研究生论文:
Rao Y (1991) Molecular and genetic analyses of cellularcommunication in Drosophila neurogenesis. University of California, SanFrancisco, California.
注2:对于依赖性分化和自主分化,摩尔根总结为:“分裂的卵之不同部分协同工作、既相互作用形成结果,或分裂导致卵的一部分与另一部分分开而独立发育?”(Morgan, 1897)。依赖性和自主分化也对应于当时所谓调节性(regulatory)发育和镶嵌性(mosaic)发育,调节性意味着早期的细胞的命运并未固定、可以变化,镶嵌性意味着胚胎早期特定的前体细胞(或区域)能、也只能发育为成体特定的细胞(或组织)。现代的术语归纳为:外源性(extrinsic)因素和内源性(intrinsic)因素在胚胎细胞发育过程中起作用。
注3:德文entwicklungsmechanik英译常为developmental mechanics、中译“发育力学”,但Sander认为Roux用mechanik应该是取康德的理解,为“自然的因果”(Sander,1991a),本文采纳此说而译为“发育成因学”。
注4:胚胎和解剖学家GustavJacob Born为物理学家Max Born(1882-1970)之父。
注5:华盛顿大学的德裔美国科学家ViktorHamburger于1988年很好地总结了Spemann的工作,但未提到Spemann and Mangold(1924)原文引用的Lewis(1907)。
注6:Hilde Mangold(图17右)于1921年23岁生日当天与Otto Mangold结婚,1923年2月提交研究生论文,其后与Otto合作做过更多实验,包括怀孕期间,实验记录于1923年12月儿子出生后停止。不幸的是,1924年9月她在丈夫的农村老家因使用酒精炉为儿子准备食物,酒精外溢烧伤过重而去世(Fässlerand Sander,1996),她的独子在二战中去世,丈夫Otto Mangold曾接任Spemann的职位,但二战后作为纳粹支持者被冷藏。
Spemann不仅以自己的发现、而且通过一批学生(包括移民美国的Holtfretter和Hamburger),推动了欧洲和美国的发育生物学研究。他于1935年获诺贝尔奖(图17),1941年去世。
进一步阅读:
Aristotle (384-322, BC) De Generatione Animalium, translated byA Platt, Oxford: Clarendon Press, 1910.
Bautzmann H, Holtfreter J, SpemannH, Mangold O (1932) Versuche zur analyse der induktionsmittel in derembryonalentwicklung. Naturwissenschaften20:971-974.
Beetschen JC (1996) How did urodeleembryos come into prominence as a model system? International Journal of Developmental Biology 40:629-636.
Born G (1896) Über verwachsungsversuche mit amphibienlarven. Archiv für Entwicklungsmechanikder Organismen4:349-465.
Driesch H (1892) Entwicklungsmechanische Studien I. Der Werthder beiden ersten Furchungszellen in der Echinodermentwicklung. ExperimentelleErzeugen von Theil- und Doppelbildung. Zeitschriftfür wissenschafliche Zoologie 53:160-178.
Endres H (1896) Anstichversuche an eiern von Rana fusca. 2.Theil:erganzun durch anstichversuche an eiern von Rana esculenta, sowietheoretische folgerungen aus beiden versuchsreihen. Archiv für Entwicklungsmechanik der Organismen 2:517-543.
Fäler PE (1996). Hans Spemann (1869-1941)and the Freiburg school of embryology. InternationalJournal of Developmental Biology 40:49-57.
Fässler PE and Sander K (1996) Hilde Mangold (1898-1924) andSpemann’s organizer: achievement and tragedy. Roux’s Archive of Developmental Biology 205:323-332.
Fischer FG, Wehmeier E (1933) Zur kenntnis der inductionsmittelin der embryonalentwicklung. Naturwissenschaften21:518.
Fischer J-L (1990) Experimental embryology in France(1887-1936). International Journal ofDevelopmental Biology 34:11-23.
Gerhart J, Danilchik M, Doniach T, Roberts S, Rowning B, StewartR (1989) Intracellular signaling centers: their establishment and modes ofaction. Development 107:37-51.
Gimlich RL and Cooke J (1983) Celllineage and the induction of second nervous systems in amphibian development. Nature 306:471-473.
Gimlich RL, Gerhart JC (1984)Early cellular interactions promote embryonic axis formation in Xenopus laevis.Developmental Biology 104:117-130.
Grainger RM, Henry JJ, and Henderson RA (1988). Reinvestigationof the role of the optic vesicle in embryonic lens induction. Development 102:517-526.
Hamburger V (1988) The heritage ofexperimental embryology: Hans Spemann and the organizer. Oxford UniversityPress, New York.
Hamburger V (1997) Wilhelm Roux:visionary with a blind spot. Journal of the History of Biology 30:229–38.
Harrison RG (1898) The growth andregeneration of the tail of the frog larva. Studied with the aid of Born’smethod of grafting. Archivfür Entwicklungsmechanik der Organismen 7:430-485.
Hemmati-Brivanlou A, Melton DA(1994) Inhibition of activin receptor signaling promotes neuralization inXenopus. Cell 77:273-281.
Herlitzka A (1895) Contributo allostudio della capacita evolutiva dei due primi blastomeric nell’uovo di triton(Triton cristatus). Archivfür Entwicklungsmechanik der Organismen 2:352-369.
Hertwig O (1893) Über den werth der ersten furchungszellen fur dieorganbildun des embroys. Experimentelle studien am frosch-und Tritonei. Archiv für Mikroskopische Anatomie 42:662-07.
Hill KA (1985) Hartsoeker’shomunculus: a corrective note. Journal ofthe History of the Behavioral Sciences 21:178-179.
Jacobson (1982) Origins of thenervous system in amphibians. In Neuronal Development (ed. Spitzer NC), pp.45-99, Plenum Press, New York.
Jacobson M (1983) Clonalorganization of the central nervous system of the frog. III. Clones stemmingfrom individual blastomeres of the 128-, 256-, and 512-cell stages. Journal of Neuroscience 3:1019-1038.
Kimelman D, Kirschner M (1987)Synergistic induction of mesoderm by FGF and TGF-β and the identification of anmRNA coding for FGF in the early xenopus embryo. Cell 51:869-877.
Lamb TM, Knecht AK, Smith WC,Stachel SE, Economides AN, Stahl N, Yancopopous GD, Harland RM (1993) Neuralinduction by the secreted polypeptide noggin. Science 262:713-718.
Lewis WH (1904) Experimentalstudies on the development of the eye in amphibian. I. On the origin of thelens in Rana palustris. American Journalof Anatomy 3:505-536.
Lewis WH (1906) Experimentalstudies on the development of the eye in amphibian. III. On the origin anddifferentiation of the lens. AmericanJournal of Anatomy 6:473-509.
Lewis WH (1907) Transplantation ofthe lips of the blastopore in Rana palustris. American Journal of Anatomy 7:137-143.
McMahon AP and Moon RT (1989)Ectopic expression of the proto-oncogene int-1 in Xenupus embryos leadsto duplication of the embryonic axis. Cell 58:1075-1084.
Maienschein J (2008) Epigenesis andpreformationism. Zalta EN (ed.), the Stanford Encyclopedia of Philosophy.
Meyer AW (1931) Essays on the History of Embryology. Palo Alto:Stanford University Press.
Morgan TH (1895) Half embryos and whole embryos from one of thefirst two blastomeres of the frog’s egg. AnatomischeAnzeiger 10:623-628.
Morgan TH (1897) The development of the frog’s egg: anintroduction to experimental embryology. MacMillan, New York.
Morgan TH (1905) The relation between normal and abnormaldevelopment off the embryo of the frog: X. A re-examination of the early stagesof normal development from the point of view of the results of abnormaldevelopment. Archiv für Entwicklungsmechanikder Organismen 19:588-614.
Needham J (1959) A History of Embryology. New York: Abelard-Schuman.
Needham J, Waddington CH, Needham DM (1934) Physico-chemicalexperiments on the amphibian organizer. Proceedingsof the Royal Society (London) B 114:393-422.
Nieuwkoop PD (1969a): The formation of the mesoderm in urodeleanamphibians. I. Induction by the endoderm. WilhelmRoux’ Archive of Developmental Biology 162:341-373.
Nieuwkoop PD (1969b): The formation of the mesoderm in urodeleanamphibians. II. The origin of the dorso-ventral polarity of the mesoderm. Wilhelm Roux’ Archive of DevelopmentalBiology 163:298-315.
Nieuwkoop PD (1970): The formation of the mesoderm in urodeleanamphibians. III. The vegetalizing action of the Li ion. Wilhelm Roux’ Archive of Developmental Biology 166:105-123.
Nieuwkoop PD, Ubbels GA (1972): The formation of the mesoderm inurodelean amphibians. IV. Qualitative evidence for the purely “ectodermal” originof the entire mesoderm and of the pharyngeal endoderm. Wilhelm Roux’ Archive of Developmental Biology 169:185-199.
Piccolo S, Sasai Y, Lu B, De Robertis EM (1996): Dorsaoventral patterning in Xenopus: inhibitionof ventral signals by direct binding of chordin to BMP-4. Cell86:589–598.
Rebagliati MR, Weeks DL, Harvey RP, Melton DA (1985).Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell42:769–777.
Roux W (1888) Beiträge zur Entwickelungsmechanik des Embryo.Über die künstliche Hervorbringung halber Embryonen durch Zerstörung einer derbeiden ersten Furchungskugeln, sowie über die Nachentwickelung (Postgeneration)der fehlenden Körperhälfte. VirchowsArchiv 114:113-153. Translated in Benjamin William and Jane M. Oppenheimer (Eds),Foundations of Experimental Embryology, pp. 2-37, 1964, Hafner, New York.
Roux W (1892) Ziele und wege der entwickelungsmechanik. Ergeb Anat Entwickelungsgesch 2:415-445.
Sander K (1991a) Wilhelm Roux and his programme fordevelopmental biology. Roux’s Archive ofDevelopmental Biology 200:1-3.
Sander K (1991b) “mosaic work” and “assimilating effects” inembryogenesis: Wilhelm Roux’s conclusions after disabling frog blastomeres. Roux’s Archive of Developmental Biology200:237-239.
Sander K (1991c) When seeing is believing: Wilhelm Roux’smisconceived fate map. Roux’s Archive ofDevelopmental Biology 200:177-179.
Sander K (1992) Shaking a concept: Hans Driesch and the variedfates of sea urchin blastomeres. Roux’sArchive of Developmental Biology 201:265-267.
Sander Kand Fässler PE (2001)Introducing the Spemann-Mangold organizer: experiments and insights thatgenerated a key concept in developmental biology. International Journal ofDevelopmental Biology 45:1-11.
Sasai Y, Lu B, Steinbeisser H, Geissert D, Gont LK, De RobertisEM (1994) Xenopus chordin: a novel dorsalizing factor activated byorganizer-specific homeobox genes. Cell 79:779-790.
Smith JC (1987). A mesoderm-inducing factor is produced byXenopus cell line. Development 99:3-14.
Smith JC and Slack JMW (1983) Dorsalizationand neural induction:properties of the organizer in X. Laevis. Journal of Embryology and Experimental Morphology78:299 -317.
Smith WC, Harland RM (1992)Expression cloning of noggin, a new dorsalizing factor localized to the Spemannorganizer in Xenopus embryos. Cell70:829-840.
Spemann H (1901) Entwickelungsphysiologische Studien amTriton-Ei. Archiv für Entwicklungsmechanikder Organismen 12:224-264.
Spemann H (1901b) Über korrelationen in der entwicklung desauges. Verhandl Anat Ges 15:61-79.
Spemann H (1902) Entwickelungsphsiologische Studien amTriton-Ei. II. Archiv für Entwicklungsmechanikder Organismen 15:448-534.
Spemann H (1903) Entwicklungsphysiologische studien amTriton-Ei. Archiv für Entwicklungsmechanikder Organismen 16:551-631.
Spemann H (1916) Über transplantationen an amphibienembryonen imgastrulastadium. Sitz Ber. Gesell NaturfFreunde zu Berlin 9:306-320.
Spemann H (1918) Über die Determination der ersten Organanlagendes Amphibienembryo I-IV. Archiv für Entwicklungsmechanikder Organismen 43:448-555.
Spemann H (1919) Experimentelle forschungen zum determinations-und individualitäts-problem. Naturwissenschaften7:581-591.
Spemann H (1920). Mikrochirurgische Operationstechnik. Abderhaldens Handb. d. bio Arbeitsmethoden2:1-30
Spemann H (1921). Über die Erzeugung tierischer Chimären durchheteroplastische embryonale Transplantation zwischen Triton cristatus und Tritontaeniatus. Archiv für Entwicklungsmechanikder Organismen 48:533-570.
Spemann (1924) Vererbung und entwicklungsmechanik. Naturwissenschaften 12:65-79.
Spemann H and Mangold H (1924). Über Induktion vonEmbryonalanlagen durch, Implantation artfremder Organisatoren. Archive fürMikroskopische Anatomie Entwicklungsmechanik 100:599-638.(translated as Spemann H and Mangold H (2001). Induction of embryonic primordiaby implantation of organizers from a different species. InternationalJournal of Developmental Biology 45:13-38).
Vogt W (1923) Morphologiesche und physiologische fragen derprimitiventwicklung, versuche zu ihrer lösung mittels vitaler färbmarkierung. SitzungsberGes Morph Physiol München 35:22-32.
Vogt W (1929) Gestaltungsanalyse am amphibienkeim mit örtlichervitalfärbung. II. Gastrulation und mesodermbildung bei Urodelen und Anuren. ArchEntw Mech Org 120:384-706.
Waddington CH, Needham J, Needham DM (1933) Beobachtungen überdie physikalisch-chemisch natur des organisators. Naturwissenschaften 21:771-772.
Waddington CH (1942) The epigenotype. Endeavour 1:18-20.
Waddington CH (1952) Epigenetics of Birds. Cambridge UniversityPress.
Weeks DL, Melton DA (1987). A maternal mRNA localized to thevegetal hemisphere in Xenopus eggs codes for a growth factor related to TGF-.Cell 51:861-777.
Zimmerman LB, De Jesús-Escobar JM, Harland RM (1996): TheSpemann organizer signal noggin binds and inactivates bone morphogeneticprotein 4. Cell 86:599-606.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}